Famiglia : Ornithorhynchidae
Testo © del DrSc Claudio Lanzoni

Il becco dell’Ornithorhynchus anatinus è ricco di terminazioni nervose © Giuseppe Mazza
Caratteristiche morfologiche, anatomiche e fisiologiche.
L’Ornitorinco (Ornithorhynchus anatinus Shaw, 1799) ha corpo tozzo e piatto e folta pelliccia impermeabile, di colore marrone scuro sul dorso e giallastra sul ventre, su tutto il corpo, tranne nei piedi e sul becco. La pelliccia copre anche occhi e orecchie (che mancano di padiglione auricolare).
La coda è costituita principalmente da tessuto grasso che viene utilizzato per immagazzinare le riserve, sfruttate dall’animale durante l’inverno o in periodi con scarsità di cibo. Essa, lunga fino a 10-15 cm, ricorda quella di un castoro, ma ne differisce sia per forma che per impiego. La coda del castoro, infatti, piatta e larga, viene usata per avanzare nell’acqua, mentre l’ornitorinco, utilizzando per la propulsione in acqua le zampe anteriori, usa coda e arti posteriori solo per cambiare direzione.
Le zampe sono corte con cinque dita fornite di lunghi artigli e le anteriori sono palmate. L’andatura sulla terra ferma è rettiliana (le zampe sono poste ai lati del corpo).
I maschi adulti sono mediamente un terzo più grandi delle femmine (circa 50 cm di lunghezza e 2 kg di peso). Essi presentano, su ognuna delle zampe posteriori e più precisamente sul calcagno uno sperone corneo lungo circa 1,5 cm. Questo sperone è cavo ed è collegato, attraverso un condotto, ad una ghiandola velenosa. Le femmine giovani presentano una versione rudimentale di questa ghiandola, che viene tuttavia persa nel primo anno di vita. Anche l’echidna possiede speroni rudimentali e ghiandole velenifere (crurali) non funzionanti.
Il becco dell’ornitorinco, provvisto di due narici, è piuttosto flessibile (simile a quello di un’anatra), morbido, gommoso e molto sensibile al tatto. Questa caratteristica è dovuta alla presenza di numerose terminazioni nervose. Il becco viene utilizzato dall’animale per cercare il cibo ed orientarsi sott’acqua. Questo animale è privo di denti (che però sono presenti nei neonati) e tritura il cibo grazie a delle placche taglienti all’interno della bocca.
L’ornitorinco è provvisto di un sistema sensoriale specializzato per percepire la presenza di prede sott’acqua. Esso si basa sulla rilevazione di campi elettrici generati dalle attività metaboliche delle prede. È il loro becco che è fornito di una serie di elettrorecettori localizzati in file rostro-caudali e di meccanorecettori uniformemente distribuiti lungo tutta la sua lunghezza. L’area elettrosensoriale della corteccia cerebrale è contenuta all’interno dell’area somato-sensoriale tattile e alcune cellule corticali ricevono dei segnali sia dagli elettrorecettori che dai meccanorecettori, suggerendo una stretta associazione tra il senso tattile e quello elettrico.

Le zampe anteriori, palmate, sono uguali in entrambi i sessi © Giuseppe Mazza
Come abbiamo precedentemente detto, durante la ricerca del cibo, l’ornitorinco utilizza prevalentemente il becco col quale scandaglia il fondo dei corsi d’acqua. È possibile che gli elettrorecettori vengano utilizzati anche per distinguere oggetti “animati” da oggetti “inanimati”.
Tuttavia, il sistema di elettrolocalizzazione dell’ornitorinco presenta ancora molti lati oscuri e richiederebbe indagini approfondite. Rispetto agli altri mammiferi ha un metabolismo piuttosto basso e la sua temperatura corporea media risulta inferiore a quella degli altri (32 °C).
Ecologia e comportamento
Gli ornitorinchi sono animali notturni semi- acquatici, ottimi nuotatori, che vivono in fiumi, laghi e torrenti di quasi tutta l’Australia e Tasmania, anche se sono molto più diffusi nel sud est del continente australiano dove meno frequentemente si verificano piene di fiumi (che provocano danni ai loro nidi) o più rara è la presenza di coccodrilli (per i quali sono facili prede).
Si tratta di animali piuttosto solitari, con specifici territori in cui vivono e si nutrono. Il territorio di un ornitorinco può essere sovrapposto con quello di un altro, ma non sembra che vi siano scontri per il suo controllo. Egli costruisce due tipi diversi di tane: tane per vivere e tane-nido. Mentre le prime sono utilizzate da entrambi i sessi per mangiare e dormire, le seconde, più complesse, sono utilizzate dalle madri per allevare i cuccioli.
Le tane-nido possono essere fino a 7 metri di altezza e 18 metri di lunghezza ed sono provviste di entrate sia sott’acqua che in superficie. Durante la stagione degli amori la femmina chiude le entrate della tana ogni volta che entra ed esce, i piccoli nati rimangono nel nido per circa tre mesi. Le tane-nido sarebbero provviste di un ingresso in superficie sia per favorire l’ingresso dell’ossigeno che per proteggere i cuccioli dalle piene.
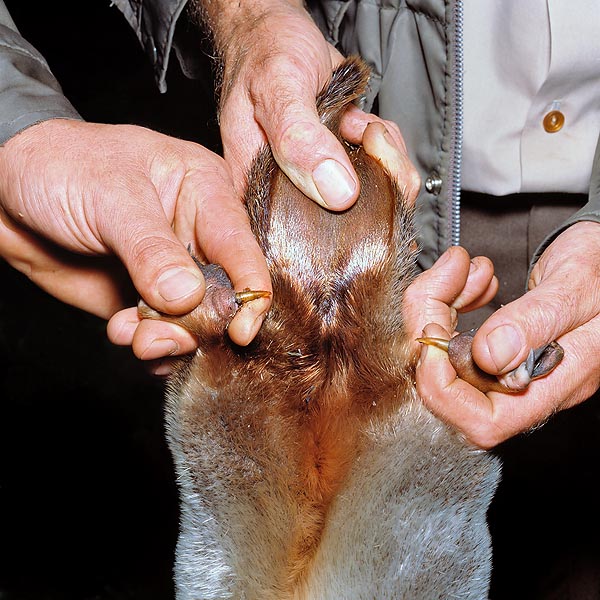
Quelle posteriori del maschio possiedono in più uno sperone velenoso © Giuseppe Mazza
L’ornitorinco è solito uscire di notte alla ricerca di cibo. È un carnivoro e della sua dieta fanno parte invertebrati (crostacei, lombrichi, insetti), uova di pesci e anfibi e piccoli vertebrati (pesci, rane, girini).
Durante l’estate si nutre molto più abbondantemente che in inverno, accumulando le riserve in eccesso nella coda, sotto forma di grasso.
La ricerca del cibo avviene durante le immersioni nei fondali dei fiumi utilizzando il capo ed il becco per smuovere il fango.
Le immersioni durano circa 40 secondi e durante questo periodo l’ornitorinco utilizza, per localizzare le prede, sia il tatto che la sua capacità di percepire variazioni del campo elettrico (grazie alle terminazioni nervose del becco del quale abbiamo già fatto cenno).
Una volta catturata la preda, essa viene trasferita nella tasca guanciale, localizzata dietro il becco. L’ornitorinco raggiunge poi la superficie e il cibo viene spostato nella bocca, dove viene sminuzzato dalle placche trituratrici situate nella mascella e nella mandibola.
Ai lati della mascella vi sono delle punte dentellate che vengono utilizzate per espellere i rifiuti (gusci, fango, ecc.).
Riproduzione
La maturità sessuale viene raggiunta attorno ai due anni. Prima di questo periodo i maschi non producono sperma e le femmine producono saltuariamente uova. La stagione degli amori è compresa tra i mesi di Luglio e Agosto. Il corteggiamento inizia in acqua. La femmina di solito prende l’iniziativa, i due nuotano molto vicini e l’accoppiamento avviene pure in acqua. Il periodo di gestazione è di circa 2-3 settimane. La femmina non partorisce piccoli vivi, ma depone uova in un nido. Le uova deposte dalla femmina sono di solito due. Esse vengono incubate per circa 10 giorni dalla madre in una camera localizzata in fondo alla tana-nido.
I cuccioli, privi di pelo, alla nascita misurano circa 18 mm; essi rimangono nella tana, nutrendosi del latte materno, per 3 o 4 mesi. Anche una volta usciti, continuano a nutrirsi del latte della madre per un certo periodo, fino a quando non sono in grado di procurarsi il cibo autonomamente. Il corpo materno produce una specie di “latte” (in cui è assente il lattosio).
Esso viene somministrato ai cuccioli attraverso due strutture, simili a capezzoli, situate sull’addome e circondate da pelo. I nuovi nati, premendo su queste zone, provocano la fuoriuscita del “latte” che cola lungo i peli e poi leccano.
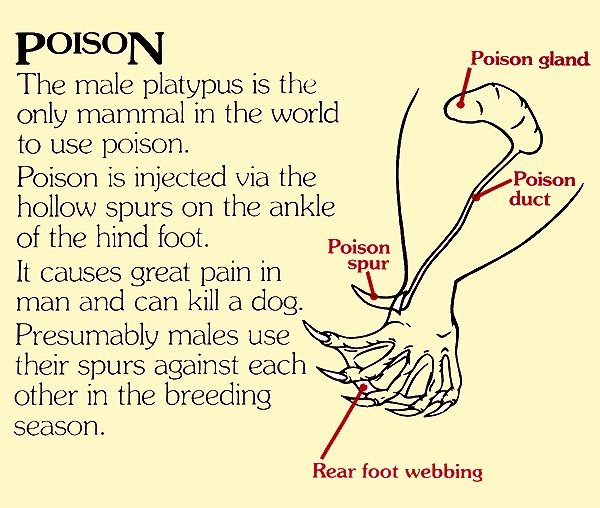
Come indica questo pannello del Healesville Sanctuary lo sperone è collegato ad una ghiandola velenifera
Anche l’analisi genetica dell’ornitorinco ha documentato che l’animale è un mosaico di caratteristiche più complesso di quanto si immaginasse (Grützner et al., 2003).
La specie si sarebbe separata dall’antenato comune che ha con la specie umana circa 170 milioni di anni fa. Recentemente si è scoperto che esso possiede 10 cromosomi sessuali (10 X nella femmina e 5 X e 5 Y nel maschio) (Waters et al., 2005).
I ricercatori hanno cercato nel genoma dell’ Ornithorhynchus sia le sequenze di DNA che appartengono unicamente ai Monotremi sia quelle presenti in altri Vertebrati scoprendo che alcune sue sequenze genetiche sono più simili a quelle che si trovano negli Uccelli che a quelle presenti in altri Mammiferi. (Grützner & Graves, 2004, Rens et al., 2007).
Il cromosoma DMRT1, ad esempio, da tempo all’attenzione dei genetisti perché sembra determini il sesso negli uccelli (sistema WZ) si ritrova quasi uguale anche nell’ornitorinco (Veyrunes et al., 2008).
Ciò ha fatto ipotizzare che la comparsa dei sessi non sia avvenuta separatamente in Uccelli e Mammiferi, ma che una parte di questa differenziazione sia comune.
L’apparato velenifero dell’Ornitorinco : struttura e funzione
L’apparato velenifero o sistema crurale degli ornitorinchi è ristretto ai maschi. Esso consiste di un paio di speroni cornei cavi, mobili, uno su ognuna delle zampe posteriori, localizzati a livello del calcagno. Ogni sperone è circondato da muscolatura ed è attaccato, con la sua base, ad una articolazione che gli consente di essere eretto, quando necessario. Alla base dello sperone è situato un serbatoio da cui si diparte un sottile condotto che lo percorre. Il canale centrale dello sperone di un ornitorinco maschio adulto ha un diametro di circa 0,2 mm (Whittington et al., 2009).
Larghi condotti, attraversano i bicipiti, connettono i serbatoi alle ghiandole velenose (alveolari, a forma di rene). Quando il maschio attacca un rivale erige i propri speroni e li pianta nelle carni della vittima; il veleno viene inoculato tramite ripetute iniezioni e, in ogni attacco, possono essere iniettati fino a 4 ml di veleno (Whittington et al., 2009). In passato molte sono state le ipotesi circa la funzione dello sperone e delle secrezioni delle ghiandole femorali.

L'andatura sulla terra ferma è impacciata e rettiliana © Giuseppe Mazza
Gli speroni possono essere utilizzati a scopo difensivo per contrastare potenziali predatori (come coccodrilli, rapaci, diavoli della Tasmania, ecc…) ma, durante la stagione degli amori, rappresentano anche un ottimo sistema offensivo per avere la meglio su altri maschi della stessa specie. L’ipotesi prevalente, già formulata da Martin e Tidswell (1895), era che, essendo presente solamente negli individui adulti di sesso maschile, gli speroni e le ghiandole del veleno dovevano essere connessi ad operazioni di selezione sessuale.
La constatazione che l’ornitorinco ha pochi nemici naturali è la dimostrazione che esiste una relazione stretta tra attività secretoria delle ghiandole femorali e riproduzione; durante la stagione riproduttiva, infatti, la ghiandola del veleno aumenta di dimensioni, aumenta la quantità di veleno prodotto e parallelamente aumentano le dimensioni del testicolo (Temple-Smith,1973; Grant e Temple-Smith, 1998; Torres e Kuchel, 2000): ciò avallerebbe l’idea che il sistema crurale si sia evoluto per avere una funzione riproduttiva.
L’avvelenamento dei rivali conferirebbe un vantaggio ai maschi dominanti: l’iniezione del veleno, causando dolore e impedendo la locomozione, permetterebbe al maschio dominante di accoppiarsi liberamente, senza aver bisogno di combattere. L’avvelenamento di un ornitorinco da parte di un altro maschio solitamente non si rivela fatale. Le manifestazioni cliniche dell’avvelenamento da ornitorinco sono state descritte in pochi lavori.
I primi casi documentati di avvelenamento umano risalgono alla fine del 1800 (Martin e Tidswell, 1895), ma la descrizione del primo caso clinico risale solo al 1992 (Fenner et al., 1992). In questo lavoro vengono riportati i principali sintomi dell’avvelenamento che sono edema ed immediato, acuto dolore che non si placa neppure con la morfina e che persiste per diversi mesi (Fenner et al., 1992). Test ematologici di laboratorio hanno rivelato un aumento della velocità di eritrosedimentazione, indicando una probabile coagulopatia, non sembrano però esserci evidenze di neurotossicità, miotossicità o effetti necrotizzanti. Non sono mai stati riportati casi fatali per l’uomo. Sintomi simili (dolore ed edema) sono stati descritti anche da autori successivi (Tonkin e Negrine, 1994).

In acqua invece è agilissimo e percepisce i campi elettrici generati dalle prede © Giuseppe Mazza
La conoscenza delle sostanze che compongono il veleno del platipo è ancora scarsa. Poche sono le ricerche condotte finora in questo campo e ricerche addizionali sono difficoltose da compiere a causa dei problemi legati all’ottenimento del veleno e di campioni tissutali.
Il veleno dell’ornitorinco è una sostanza piuttosto complessa. L’inusuale natura dei sintomi di avvelenamento farebbe pensare ad una miscela complessa di molti differenti peptidi e proteine, alcune delle quali ancora sconosciute (de Plater et al., 1995; Whittington e Belov, 2007).
I primi esperimenti condotti sulla secrezione delle ghiandole crurali hanno dimostrato che il veleno provocava, nei conigli, per iniezione sottocutanea, importante edema e rapida diminuzione della pressione arteriosa (P.A.), difficoltà respiratorie e morte, se iniettata per via venosa. Negli anni’90, la disponibilità di nuove tecniche per la caratterizzazione delle proteine ha permesso di identificare con precisione alcuni componenti del veleno dell’ornitorinco. In alcuni studi è stato dimostrato l’effetto del veleno sulla muscolatura liscia, facendo ipotizzare la presenza, al suo interno, di un peptide natriuretico (de Plater et al., 1995; 1998 a, b). I peptici natriuretici sono una famiglia di peptidi che si sono altamente conservati nelle diverse specie e che appaiono controllare la pressione sanguigna.
È stato scoperto di recente che il veleno consiste di almeno 19 differenti frazioni peptidiche (Whittington e Belov, 2007) insieme a componenti non proteiche (probabilmente prostaglandine). Attualmente, però, solo tre peptidi sono stati identificati e sequenziali, anche se le loro esatte funzioni non sono ancora conosciute con precisione. Essi sono: peptidi natriuretici di tipo C (CNPs); peptidi defensivo-simili (DLPs); nerve growth factor (NGFs).
Sinonimi: Ornithorhynchus brevirostris Ogilby, 1832; Ornithorhynchus crispus MacGillivray, 1827; Ornithorhynchus fuscus Péron, 1807; Ornithorhynchus laevis MacGillivray, 1827; Ornithorhynchus novaehollandiae Lacépède, 1800;
Ornithorhynchus paradoxus Blumenbach, 1800; Ornithorhynchus phoxinus Thomas, 1923; Ornithorhynchus rufus Péron, 1807; Ornithorhynchus triton Thomas, 1923.
→ Per nozioni generali sui MAMMIFERI cliccare qui.
→ Per apprezzare la BIODIVERSITÀ dei MAMMIFERI cliccare qui.