Phylum : Amoebozoa

Text © Prof. Giorgio Venturini

English translation by Mario Beltramini

Amoeba proteus in vital stain, with the Neutral Red concentrating in the lysosomes © Dr. Giuseppe Mazza
Amoeba proteus Pallas, 1766, is the most known species of a genus of protists previously inserted in the phylum Rhizopoda, but presently, on the base of molecular data, classified as phylum Amoebozoa, belonging to the Kingdom of the protists. We have however to keep in mind that the classification of the protists is presently much debated.
Most of the species are free-living in the soil, in the mud or in the water, where they nourish of bacteria, small protists and other unicellular organisms; only few species are parasitic.
Amoeba proteus is the best studied species: it is an unicellular organism of about 250 micron (μm) of diametre, that can reach even the 400-500 micron, able to emit lobe-shaped cytoplasmic extensions, the pseudopodia, utilized for the locomotion and the capture of the preys. The genus Chaos, cognate with Amoeba, includes the largest known amoebas, among which we recall Chaos carolinense that may reach the 5 mm.
Observed under the microscope fresh and without colourations the amoeba looks like an irregularly shaped structure, devoid of colouration and translucent, animated by continuous movements that consist in changhe of shape, emission of pseudopodia and continuous flow of cytoplasmic granular material. Inside it, besides several granules we observe a nucleus and a large roundish vacuole. Contrary of to the genus Chaos whose members are plurinuclear, Amoeba proteus is uninuclear.
The habitat of the amoeba consists essentially in the mud on the bottom of freshwater pools, lakes and water low flowing streams, often under the aquatic vegetation, and also in the very much humid soils.
The name of the genus Amoeba, in Italian ameba, comes from the Greek “amoibé” (ἀμοιβή) = change; Proteus in the Greek mythology is the name of a sea-god, called also the “Old Man of the Sea”, able to transform assuming the appearance of any animal or plant, and also of objects like water or fire.
The oldest description of an amoeba is due to August Johann Rösel von Rosenhof, Austrian nobleman, painter of miniatures, naturalist and entomologist who in 1755 has baptized it “Der kleine Proteus” (“the little Proteus”). Thanks to his ability as painter this scholar has left us beautiful pictures of the amoeba and of many other organisms.
Historically, the microscopists have subdivided the cytoplasm of the amoeba in two parts, an outer “ectoplasm” little granular and in contact with the cellular membrane and an inner granular “endoplasm” in its turn distinguishable in a more viscose portion, called plasmagel, and one less viscose, called plasmasol. Actually, these subdivisions reflect different functional conditions of the cytoplasm and are continuously variable.
Locomotion
To understand the locomotion of an amoeba we can think to our step, that involves the forward projection of one leg and the following contact of the foot with the soil. The foot, now well placed on the ground serves as anchorage for pulling forward the body thanks to the contraction of the muscles of the hip whilst the other foot, staying previously on the soil, raises. Also in the amoeba the principle is the same: forward projection of a pseudopod, anchorage of the same to the substratum and traction of the cell body forward, while the previous anchorages detach. We have now to understand how forms a pseudopod, how it elongates and anchor to the substratum and how occurs the traction forward of the cell body.
In order to explain the movements of the amoeba and of other cells endowed of amoeboid movements have been proposed in the time various theories that usually evoke the changes of consistency of the various parts of the cytoplasm that alternatively pass from a status of sol to one of gel (that is, from liquid to gelatinous and vice versa), allowing a sort of squeezing of the cytoplasm towards the front of advancement. In these theories is not clear what are the mechanisms causing these changes of viscosity, how they are directed, how actually they generate a movement and which are the sources of energy to accomplish this work.
Most recently it was examined the role of some proteins present in the cytoplasm of the amoeba and well known in the pluricellular organisms, that form a true and real flexible intracellular skeleton (cytoskeleton), able to lengthen and shorten actively besides to act as tracks where can move other proteins, real molecular motors able to drag in their turn other protein filaments. These cytoskeletal proteins and the related molecular motors stand also at the base of the movements of our muscles.
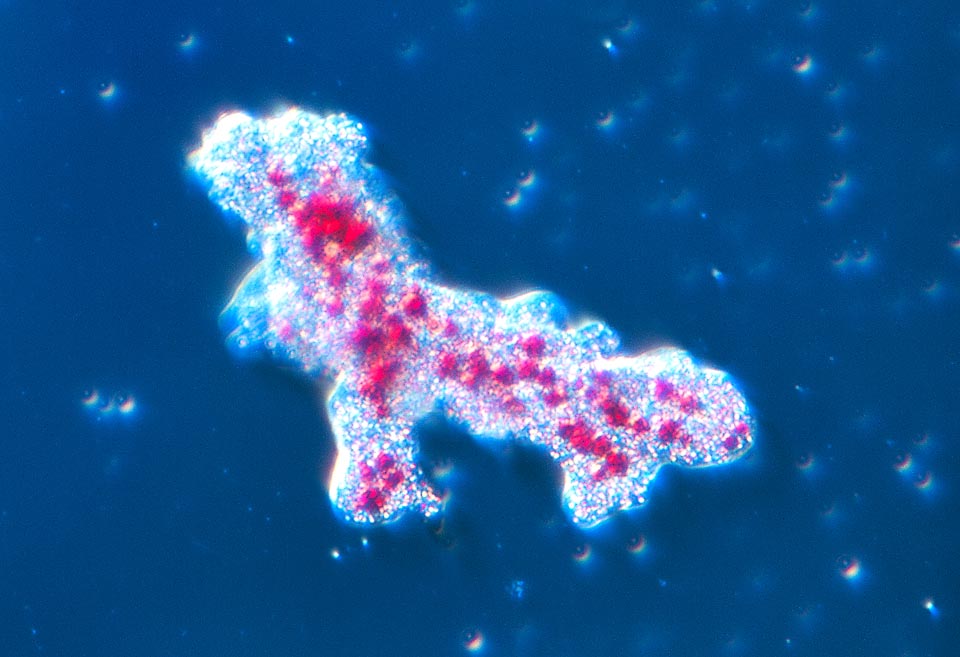
The habitat of the amoeba consists mainly in the mud on the bottom of freshwater pools, lakes and water streams with little current of all the world © Dr. Giuseppe Mazza
We try here to draw a possible mechanism (that however does not exclude the possible intervention of other phenomena). The main protein forming the cytoskeletal filaments is called actin and is an approximately globular molecule, abundant in the cytoplasm of the amoeba as well as that in our cells. The molecules of actins may join reversibly to form a filament, like a sort of pearl necklace, that can elongate or shorten by addiction or subtraction of globular molecules. It is thought that the protrusion of the pseudopodia may be at least in part generated by the elongation of the filaments of actin that should come to push ahead the membrane merging in parallel bundles.
In addition or as an alternative to this explanation has been invoked a role of the actin and of the motor proteins to it associated in producing a contractile force able to induce by pressure flows of cytoplasm towards the front of advancement. Also the change of the consistency of the cytoplasm from a more liquid status to a more solid one is consequence of the aggregation of the molecules of actin. Other proteins interact with the actin forming crossed links between the filaments and consequently generating a tridimensional grid.
For a better understanding of this phenomenon we can think to what happens when the meat broth cools down and turns into jelly: the protein molecules of the collagen, separated from each other in the boiling broth, aggregate to form filaments that create a tridimensional grid that hardens the jelly. In the example of the jelly gets form a grid with filaments casually oriented, whilst in our tissues, thanks to the intervention of the cells, the collagen aggregates to form robust parallel bands, the tendons. When the pseudopod has elongated some proteins of the membrane form anchorages to the substratum.
What are the factors that stimulate the forming of the filaments of action? What is for an amoeba “anterior” or “posterior”?
We know that the amoebae are sensitive to signals present in the environment (positive or negative tropisms), for instance, they are attracted by molecules released by sources of food: these molecules bind receptor proteins present on the membrane of the amoeba and activate a molecular response that stimulates the aggregation of the globular actin to form the filaments. It is interesting to note that the molecules stimulating the aggregation of the actin in the amoeba are the same performing the same task in our white blood cells or in many other mobile cells. It is therefore the chemical signal coming from the environment that stimulates the protrusion of the pseudopods and that determines what will be the anterior pole of the cell. When the signal disappears the pseudopods stop elongating in that direction since the filaments of actin disaggregate. The next step requires the dragging or the pushing of the cell body in direction of the pseudopod: here come into play the motor proteins (myosins) that like real and true molecular motors “walk” on the filaments of actin and determine the movement of the cytoskeletal scaffoldings of the cytoplasm in direction of the advancing extremity, whilst the old protein anchorages to the substratum detach.
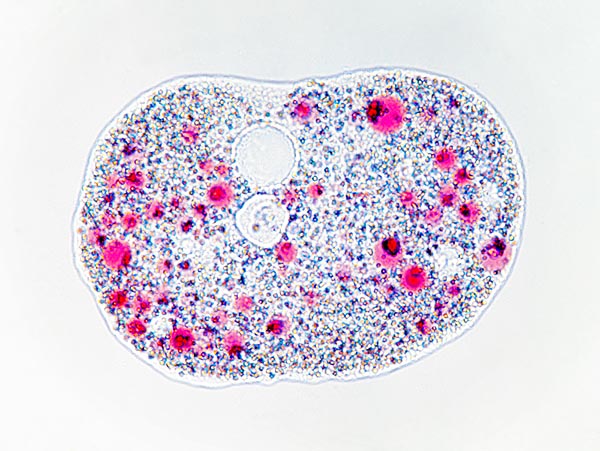
Amoeba proteus resting. At the centre on top we note the vacuole © Dr. Giuseppe Mazza
This interaction between actin and myosins is similar to that operating in our muscles. Of course, the elongation of the filaments by apposition of new sub units as well as the activity of the molecular motors like the myosin require energy that is furnished, like for most cellular phenomena requiring energy, by the well known molecule of ATP, molecule rich of energy continuously produced by the metabolism of all cells. Also fragments of cytoplasm isolated from the cell body, exhibit movements in presence of ATP.
A confirmation of the essential role of the filaments of actin in the locomotion of the amoeba comes from experiments where to the amoebae has been administered a drug (cytochalasin) which prevents the formation of the filaments of actin. In these conditions the amoeba is no longer able to emit pseudopods and to move.
It can be amazing the fact that the same molecular mechanisms operate in the movements of the amoeba, in our white blood cells or in our muscles: we can explain these similitudes reflecting on the fact that the mechanisms of the cellular movement have evolved in the distant ancestor who, almost a billion of years ago, has originated the evolutionary line of the protozoa and that this will lead to the human beings. Hence, the actin of the amoeba is the same as the human one? Surely not identical, seen that the actin of the common ancestor underwent cumulative evolutionary changes (mutations) along almost a billion of years, evolving towards the actin of the modern amoeba or towards that of the modern man, but its basic functional characteristics have remained the same.
Nutrition
The amoeba is a predatory organism that nourishes engulfing by phagocytosis other protists or bacteria. The phenomenon is made possible by its capacity of perceiving the presence of the preys thanks to chemical signals or of contact and mainly of emitting pseudopods by means of which it surrounds the microorganism and engulfs it in a membrane vesicle (phagocytosis vacuole or phagosome). Actually, for a more accurate representation of the phenomenon we must keep in mind its tridimensionality and think in this case to the pseudopods like some lips that close around the prey to insert it into the “mouth”, that is into the phagosome.
It is intuitive that the membrane surrounding the phagosoma comes from the cell membrane and therefore every phagocytosis subtracts material to the involucre. If we consider that an amoeba in one day probably carries out an average of 100 endocytoses, in each of which it internalize about the 10% of its own surface of membrane, we can calculate that every day is subtracted the equivalent of 10 cell surfaces. This is made possible by concurrent phenomena of exocytosis that return to the surface the membrane of the old phagosomes or newly formed membrane.
For understanding correctly this continous recycling of membranes in a cell and the transferring of the contents of vesicles we must think that membrane is a fluid film and the behaviour of a cell and of its systems of vesicles may be compared with that of oil drops put in a dish full of water: the drops may merge, uniting their contents or split apart to then merge again. In the same way from the membrane of the cell may detach vesicles ready to merge again, transferring their contents and their membrane in a continuous interchange.
The phagocytosed material is now sent to the digestion by fusing the phagosome with other membranous vesicles containing digestive enzymes, functionally equivalent to those present in our digestive system. Since these enzymes work correctly only in a very acidic environment, some molecular pumps present on the membrane of the phagosome actively transport hydrogen ions inside the vesicle (acidity corresponds to a high concentration of hydrogen ions). The resulting vesicle, acidic and containing the phagocytosed material and the digestive enzymes, takes the name of lysosome and carries out the function of cell digestive apparatus.
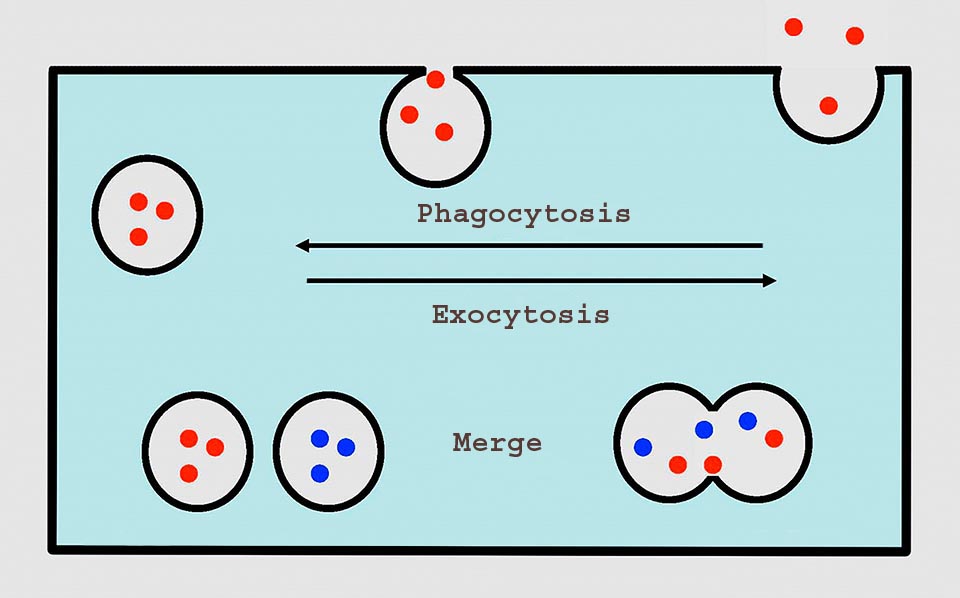
In the amoeba the membrane with the phagocytosis produces a vesicle that merges with the lysosome, after the digestion the waste products are expelled with an exocytosis that also renders the membrane the surface subtracted with the phagocytosis. Also the contractile vacuole expels its contents with an exocytosis © G. V.
When the enzymes will have completed their action the lysosome will contain digested material, useful for the amoeba, besides waste products and undigested material. At this point transporter proteins present in the membrane of the lysosome transfer into the cytoplasm the useful substances (for instance, sugars or amino acids coming from the digestion), while the lysosome gets close to the cell membrane and fuses with it, pouring out the waste products. Anyhow, never occurs the breaking of the lysosome and the pouring of all its contents into the cytoplasm; should this happen the cell would self-digest!
The comparison with our feeding is easy: the food ingested is digested by the enzymes of the stomach and the intestine, the useful substances obtained from the digestion are absorbed by the intestinal wall and transferred to the blood and the waste products are expelled with the feces. The breaking of the intestine wall and of the stomach (perforation) have extremely serious consequences.
Whilst the phagocytosis allows the amoeba to incorporate voluminous structures such as bacteria or protists, this organism may get from the environment nutrients through a process called pinocytosis (from the Greek “πινω” (pino), that means I drink), that anyway is based on principles similar to the phagocytoses, as it allows to insert in membranous vesicles water and macromolecular solutes that will be then went to the lysosome.
It should be finally remembered that the cell membrane of the amoeba holds sophisticated mechanisms of selective permeability that allow the passage of single molecules of small size, like the sugars, from the outside to the cytoplasm, without formation of vesicles.
The contractile vacuole and the water regulation
Living in freshwater environments the amoeba in order to survive must face the problem of the regulation of its own contents of water. In fact, the cytoplasm can be considered as a very concentrated solution of organic substances (such as, for instance, proteins or sugars) and inorganic, separated, thanks to the cell membrane, from a watery environment at low concentration of solutes, and therefore richer of water. Being the cell membrane permeable to the water the inner high concentration of solutes attracts inward water by osmotic effect. The inevitable result, in absence of regulation mechanims, should be that of swelling the cell and possibly of its lysis (rupturing).
The amoeba can face this problem thanks to a cellular organelle, the so-called contractile vacuole, membranous vesicle that stores inside the water in excess and then expels it out from the cell. The mechanism through which the vacuole can accumulate the water has long time been obscure, its recent explanation has allowed to unveil also other functions of this structure.
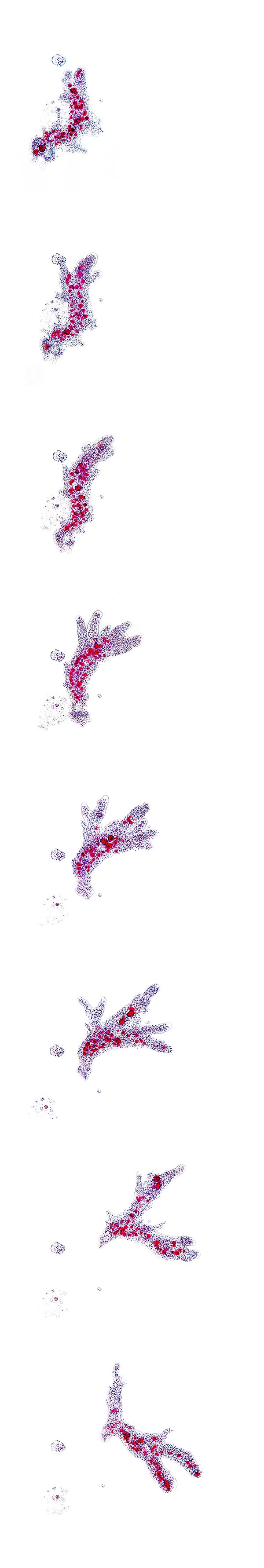
Movement sequence of an Amoeba proteus © Dr. Giuseppe Mazza
The membrane of the vacuole is equipped with channels permeable to the water and with molecular pumps able to transport actively inside it various substances, among which, mainly, hydrogen ions (protons). Thanks to this protons pumping, the interior of the vacuole has a very high concentration of ions that, by osmotic effect, attracts water from the cytoplasm. In such way the water, that due to the osmotic action of the cytoplasm had entered the cell, by osmotic effect accumulates in the vacuole (if we consider the gradient of the concentration of the solutes, this results minimal in the external environment, intermediate in the cytoplasm and maximal in the vacuole, therefore the water will transfer from outside to the cytoplasm and from this one to the vacuole).
At this point the vacuole moves to get in touch with the cell membrane and opens outwards pouring its contents of water and of solutes. Actually, on the membrane of the vacuole are present, besides the pumps for the hydrogen ions, also other transporters that accumulate inside it various substances, such as for instance metabolic wastes like carbon dioxide, urea or uric acid, that will be expelled outwards when the vacuole pours its contents out of the cell. As a consequence, the vacuole does not act only as organelle regulator of the contents of water but also as excretory organelle that eliminates the wastes of the metabolism. The expulsion of the contents of the vacuole implies that its membrane fuses with the cell membrane and therefore at this point the vacuole disappears: a new one will form “ex novo” through the production of a new vesicle.
Obviously, the transportation of hydrogen ions and of other substances inside the vacuole requires energy, and this is provided by the molecule of ATP.
The term contractile vacuole suggests a capacity of contracting actively, suggested by the observations of the microscopists that had noted its periodical changes of volume. Actually, it seems that these changes are mainly due to the water accumulation and not to phenomena of active contraction.
In the amoebae living in sea water, that do not have the problem of the swelling seen that the sea water has a saline concentration higher than that of the cytoplasm, the vacuole does not exist (these amoebae will have, conversely, the opposite problem of the dehydration).
Genetics and reproduction
As in the case of other giant amoebae, also Amoeba proteus has in its nucleus a huge number of chromosomes, about 250 pairs, and an enormous quantity of DNA, estimated in about 290 billions of bases (in the man we have 23 pairs of chromosomes and 3 billions of bases). The very high values found in Amoeba proteus come from phenomena of polyploidy, that is, the chromosomes have repeatedly duplicated in identical pairs. The cited numbers are furthermore variable during the vital cycle of the protozoan. The functional meanings of these phenomena are not clear.
Reproduction
The reproduction of the Amoeba proteus is a periodical process repeating in intervals that depend from the rate of increase. An amoeba will start dividing when reaches a sufficient size, for instance, 0,2-0,3 mm. The reproduction is asexual and usually occurs by binary fission. Before dividing, the cell retracts the pseudopods and assumes a roundish shape, therefore occurs a nuclear mitotic division followed by subdivision of the cytoplasm that happens thanks to a cleavage furrow (due to the same contractile proteins acting in the locomotion) followed by the activity of the pseudopods that drag in opposite directions of the two daughter cells. The process usually lasts 30 to 60 minutes.
They have described phenomena of multiple fission, that should occur in unfavourable conditions of poor feeding or of dehydration: the amoeba gets round and covers with a resistant and impermeable cyst that enables it to survive till when the conditions become favourable. At this point a multiple nuclear division produces many nuclei that gather at the periphery of the cell and finally separate in many new cells that get out when the cyst hydrates and bursts.
The encystment seems to happen also independently to the multiplication and represents therefore a system for overcoming periods of adverse conditions, such as the drying of the pool of water.
Some microscopists have described phenomena that lead to think to a conjugation, but the meaning of these observations is not clear.
Symbiosis and parasitism
Even if Ameba proteus is able to phagocytize and digest most bacteria, there is at least one species of bacterium, similar to the Legionella, the pathogenic agent of the Legionnaires’ disease, that is able to survive inside the phagosome, avoiding the digestion. This bacterium establishes a symbiotic relation with the amoeba which induces in both species changes that render them mutually dependent.
In some species of amoeba have been found infections by viruses characterized by exceptionally large dimensions, such as the Mimivirus or the Pandoravirus. These viruses have diametres between the 400 and the 1000 nanometres (nm, billionths of metre) and their DNA amounts to one or two millions of bases. We consider tha most viruses have dimensions of few tens of nm and their DNA has few tens of thousands of bases.
Predators of the amoeba
The amoebae are object of predation by various organims, among which nematodes, small fishes, crustaceans and mollusks, from which they can defend by secreting toxic or repellent substances.
Amoebae pathogenic for the man
The pathogenicity of the amoebae is a not completely clarified matter, since that only few strains of the potentially involved species result permanently pathogenic for the man and that in most of cases they easily lose their infectivity.
The most important genera are Naegleria and Balamuthia, that may cause serious encephalitis, Acanthamoeba, cause of meningoencephalitis and of keratitis, or Hartmannella, cause of keratitis. In addition to this, we must recall that some amoebae may be infected by bacteria pathogenic for the man and then act as disease vectors.
Surely the most important pathogenic species is however Entamoeba histolytica, agent of the intestinal amoebiasis, intestinal infection that can be extremely serious, with even fatal complications. It is estimated that 50 million of persons in the world are infected by this amoeba, especially in the tropical contries, with 50000-100000 deaths per year. In most cases the infection is asymptomatic, but in about the 10% of cases it causes forms of dysentery.
The amoebae invade the intestinal mucosa and destroys its cells, producing ulcerations, with loss of blood that can cause serious severe anemia and possible perforations with extremely serious results. The ulcerations can allow the protozoans to reach the blood stream and therefore to be transported into organs such as the liver or the brain causing very serious, even deadly, abscesses. When they reach the colon, due to the progressive dehydration of the intestinal contents, the amoebae produce a cyst that will be expelled with the feces and that represents the form able to infect other individuals. The mature cyst has four nuclei and rightly its microscopical research in the feces represents the classical diagnostic method. The cyst is extremely resistant and can survive long time in the water or in the ground.
The contagion occurs by ingestion of water or of food contaminated by the feces containing the cysts that, when they reach the intestine of the new host, transform in the trophozoite, that is in the active amoeba able to multiply and invade the mucosa. The amoebiasis is a typical example of fecal contagion infection, described by the English as “F Cycle”: feces, fingers, flies, food. The parasites present in the feces, through the hands or the flies reach the food. Numerous anedoctes are reported concerning cases of multiple contagion, among which that of a cook of a cruise vessel that, clearly paying little attention to the hygiene of his hands, should have infected all passengers! (We recall that in the Anglo-Saxon countries in the public toilets are posted notices “Now wash your hands!”).
To the genus Entamoeba belong numerous species, some of which, like Entamoeba histolytica, Entamoeba dispar, Entamoeba moshkovskii, Entamoeba polecki, Entamoeba coli, and Entamoeba hartmanni may live in the human intestine. Only Entamoeba histolytica is surely pathogenic. The differential diagnosis is based on the characteristics of the cyst, tetranuclear in Entamoeba histolytica, and mainly on immunological or molecular biology methods.
Amoeba in the popular culture
In the song “A very cellular song” of “The incredible string band” the life is described from an amoeba’s point of view: “Amoebas are very small, There’s absolutely no strife Living the timeless life, I don’t need a wife Living the timeless life. If I need a friend, I just give a wriggle split right down the middle and when I look there’s two of me both as handsome as can be. Oh, here we go slithering, here we go slithering and squelching on …”
Synonyms
Volvox proteus Pallas, 1766; Proteus diffluens O.F. Müller, 1786; Chaos diffluens (O.F. Müller, 1786) Schaeffer, 1926.