Le premier être vivant était une plante



Texte © Dr. Giuseppe Mazza

Traduction en français par Serge Forestier
Quand la vie est-elle apparue sur la Terre ?
Il semble que cela se soit produit il y a environ 4 milliards d’années. L’air, dépourvu d’oxygène, était un mélange, certainement pas sain, d’ammoniaque, d’hydrogène, de méthane, d’acide sulfurique et d’acide cyanhydrique. Mais sous l’effet de la chaleur, du rayonnement et de la foudre, certaines molécules de la “soupe primitive”, de l’eau de mer, à savoir, chargée de sucres, d’acides aminés, d’acides gras, de bases puriques et pyrimidiques, ont mystérieusement fusionné.
Ainsi sont nés les fameux 20 acides aminés constituant les “briques” de la matière vivante, les protéines, les codes génétiques et la fine membrane qui séparait le premier micro-organisme de l’environnement.
Il s’agissait, comme le suggèrent les restes fossiles, d’êtres proches des bactéries, qui se nourrissaient d’ATP (adénosine-triphosphate), une substance alors abondante dans la mer. Ils brisaient cette grosse molécule, un peu comme nous le faisons aujourd’hui dans les centrales nucléaires avec l’atome d’uranium, en tirant de l’énergie et un composé plus petit : l’ADP (adénosine-diphosphate).
Tout allait bien, mais après quelques millions d’années, l’ATP, logiquement, se fit rare.
C’était la première crise énergétique majeure, et la nature réagit par une réaction chimique opposée. L’ADP, maintenant en excès, se combina avec le glucose en fermentation grâce à d’autres bactéries, et réforma de l’ATP. Le cycle se referme donc admirablement : d’une part des micro-organismes qui déchargent la “batterie de la vie”, et d’autre part des micro-organismes qui la rechargent.
Mais comme cela se produit encore aujourd’hui avec le moût, le processus de fermentation produit des déchets : des bulles de dioxyde de carbone qui s’accumulent dans l’atmosphère, et de l’alcool qui se concentre dans la “soupe primitive”.
Après environ un milliard d’années, la terre est de nouveau en crise : le glucose se fait rare, et les êtres vivants nagent ivres et désespérés dans une mer de problèmes.
C’est la seconde crise énergétique majeure. La partie semble définitivement perdue, lorsqu’une géniale “bactérie”, il y a 3 100 millions d’années, selon les fossiles calcaires de Bulawayo en Rhodésie, invente une molécule verte extraordinaire : la chlorophylle. Elle se laisse traverser par les rayons du soleil, mais au lieu de s’échauffer comme un vulgaire éclat de verre, elle synthétise, d’une manière encore mystérieuse aujourd’hui, le matériel et l’immatériel.
La première plante est née. Un être capable de combiner l’énergie lumineuse, avec le dioxyde de carbone, l’eau et les minéraux, pour fabriquer de la matière vivante, et en particulier des sucres.
Mais même ce processus a un rejet : une substance alors considérée comme dangereuse et polluante : l’oxygène.
Il se rassemble peu à peu dans l’atmosphère, se transforme dans les couches supérieures en ozone, et protège la terre contre les rayons ultraviolets, teintant de bleu le ciel, jusqu’ici grisâtre. Un spectacle merveilleux, mais un malheur pour les pauvres bactéries anaérobies de la fermentation, qui vécurent, on l’imagine bien, cette découverte comme une “catastrophe nucléaire”.
Certaines s’adaptèrent aux nouvelles conditions environnementales, inventant la respiration, processus par lequel, avec un rendement 18 fois supérieur à la fermentation, on récupère de l’énergie à partir des sucres brûlant de l’oxygène, mais la plupart sont mortes ou se sont réfugiés dans le sous-sol, comme le feraient dans notre civilisation, les survivants d’une guerre atomique effroyable. Et nous les trouvons encore là, dans le noir, là où l’air est absent, sous la vase de la mer, des lacs et des étangs.
Mais quand vivaient ces êtres primitifs ?
Ils étaient potentiellement immortels. Ils se reproduisaient en quelques minutes, par scission, comme le font encore aujourd’hui les bactéries, et de nombreuses espèces unicellulaires, telles que les algues planctoniques, les amibes ou les infusoires.
Le processus était des plus trivial : lorsque, à force de manger, le micro-organisme devenait trop gros, il dupliquait son patrimoine génétique et se divisait en deux. Mais la “mère” (ou le “père”, parce que les sexes n’étaient pas encore nés !) ne disparaissait pas : rajeuni par le fractionnement, il/elle continuait à croître et à vivre, en se divisant éternellement.
Certes, quelques cellules mourraient accidentellement, dévorées par d’autres organismes, ou peut-être parce que la flaque d’eau s’était asséchée, mais à moins d’une catastrophe, l’individu, cette combinaison chromosomique, avait une espérance de vie illimitée.
Il va sans dire que les cellules d’un tel “super-être” sont toutes rigoureusement identiques. En effet, étant donné que de l’examen des acides aminés des organismes vivants (toujours les mêmes, présents de plus toujours sous la forme “L”), de la structure du système de synthèse protéique et de l’universalité du code génétique, on déduit que la vie sur la terre est née une seule fois, le monde, au début, devait être peuplé d’un organisme unique, omniprésent, avec des cellules séparées mais identiques.
Pouvez-vous imaginer quel ennui ? Le “M. Rossi” qui le matin, en quittant la maison, se retrouve dans la rue avec lui-même, pour se disputer avec lui-même au bureau, et puis rentrer se coucher avec lui-même !
Heureusement, à la longue, sous l’effet des rayonnements mutagènes et des erreurs de duplication des gènes, les cellules théoriquement identiques de notre “super-être” ont commencé à se différencier.
La nature conservait dans telle ou telle la “réponse” pour surmonter les différentes difficultés environnementales, et étant donné que, comme cela arrive encore aujourd’hui parmi les bactéries, les gènes, libérés dans le protoplasme, passent facilement d’un individu à l’autre, bien avant les Franciscains, le mot d’ordre de la vie fut “tout diviser, et emporter le moins possible avec soi “.
Le monde pendant des millénaires fut une énorme “foire aux gènes” dans laquelle, selon le besoin, avec une simple “collision”, les micro-organismes se les échangeaient dans un esprit fraternel.
Puis, il y a environ 2 milliards d’années, l’un d’entre eux est devenu gourmand et avare. Il a désobéi, il a décidé de ne plus donner ses propres gènes au premier venu, et de les enfermer dans un “coffre-fort”, le noyau, comprimés et ordonnés comme les livres d’une bibliothèque (aujourd’hui, une bactérie a en moyenne 1 million d’informations chromosomiques, une amibe 400 millions, et un homme 5 milliards !), pour faire face à toutes les éventualités.
Les échanges fraternels n’étaient plus possibles, mais entre-temps la nature avait conclu que si une cellule se divisait pour en faire deux, on pouvait bien imaginer le processus inverse : deux cellules, autrement dit, qui fusionnent pour n’en donner qu’une.
De cette façon, l’échange des chromosomes, des “recettes pour la vie”, était non seulement possible, mais il s’en trouvait renforcé.
Ainsi est née la sexualité, avec le corollaire indissoluble de la mort.
Deux organismes, assez similaires pour se “marier” mais avec des caractéristiques différentes, mettent en commun leurs gènes, et créent un nouvel être, quasiment unique (dans l’espèce humaine, par exemple, les combinaisons chromosomiques possibles sont au nombre de 64 000 milliards !), et différent des deux.
Le prix est très élevé, car les géniteurs ne peuvent plus se dupliquer par scission, tôt ou tard ils meurent, et leur identité et leur configuration génétique sont donc perdues pour toujours ; mais tandis que les “bactéries bourgeoises”, théoriquement immortelles, ont été coupées du monde par l’évolution, les “fraudeurs”, avec leur brassage continu de gènes, ont peu à peu créé, en se différenciant, tous les autres êtres vivants.
Il s’ensuit que la même division entre les animaux et les plantes n’est qu’une question de commodité.
La vie est un phénomène unique, et la parenté entre les deux règnes est beaucoup plus proche que nous le pensons. Tant et si bien que certaines algues unicellulaires “indécises” comme les Euglena, peuvent se comporter comme les plantes et faire de la photosynthèse s’il y a de la lumière, ou perdre la chlorophylle, et vivre comme des animaux, de substances organiques, dans l’obscurité des égouts.
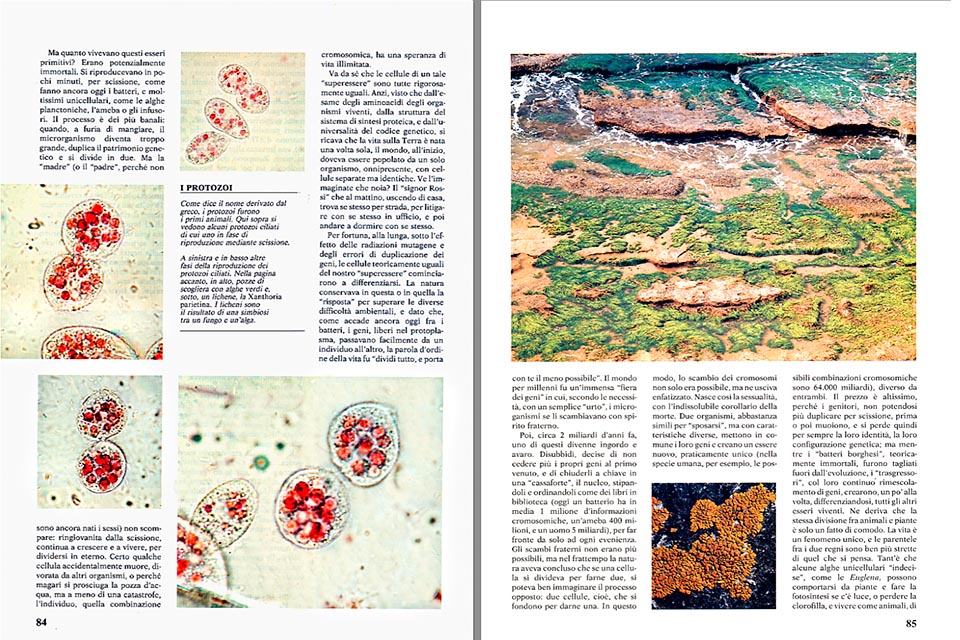
Mais revenons aux algues qui ont découvert la photosynthèse et le mariage. Certaines, comme les Diatomées, de délicats étuis siliceux, se reproduisent aujourd’hui autant par scission que par voie sexuelle. Elles sont parmi les principales composantes du phytoplancton, des “prairies sous-marines” massacrées de végétaux en suspension, qui se nourrissent directement ou indirectement, comme le fait sur la terre ferme avec les autres plantes, le monde animal environnant.
La quantité de “viande verte” produite par les deux environnements est étonnamment similaire. Dans la Manche, par exemple, où la mer avec des reflets verdâtres indiquant la présence de quantités importantes de phytoplancton, on a comptabilisé la création de 1 400 tonnes de matière végétale par an et par km², à comparer aux 1 800 tonnes de foin des prairies adjacentes.
Les mers aux eaux limpides, comme c’est le cas en revanche sur la Côte d’Azur, reflètent le bleu du ciel pour la plus grande joie des romantiques, mais aussi une pauvreté congénitale en phytoplancton, et donc malheureusement en poisson.
Mais un beau jour les algues planctoniques, lassées de se cogner le long des côtes, pensèrent que ça ne serait pas mal de les attaquer.
Dans la zone de ressac, où l’air et l’eau sont sans cesse brassés, en fait, en plus du soleil, abonde le dioxyde de carbone, “matière première” fondamentale de la photosynthèse.
Certaines algues unicellulaires ont développé dès lors en parallèle des “pieds”, filamenteux, ramifiés, ou foliacés, appelés “thalle”, pour s’ancrer solidement aux rochers. Ce fut bientôt la course pour accaparer les meilleures places.
La zone privilégiée est le niveau de la marée basse, où même pendant la pleine lune, le risque de rester sans eau est nul. Et là naturellement les algues “nobles” se sont installées, comme la majestueuse Laminaire, aux grandes lames brunes qui peuvent atteindre (Macrocystis spp.) les 150 m de longueur : le plus grand être vivant connu.
Au-dessus de cette “zone de luxe”, des organismes plus rustiques se sont installés, avec des thalles souvent riches de ballonnets flottants, comme notre Fucus ou l’incroyable Hormosira banksii australiana ; et plus haut la Laitue de mer (Ulva lactuca), avec des espèces semblables aux thalles verts de plus en plus minces, jusqu’à ce qu’elles deviennent filiformes, comme l’herbe, pour mieux résister à la déshydratation.
Au-dessous, dans les zones moins exposées au soleil, se sont implantées les algues brunes et les algues rouges, aux colorants sophistiqués associés à la chlorophylle, pour mieux exploiter, jusqu’à 200 m de profondeur, les faibles rayons solaires.
Pour se reproduire ces plantes mettent en pratique pratiquement toutes les stratégies sexuelles et asexuées connues.
Comme les Sargasses, elles peuvent recréer les “feuilles” à partir d’un fragment de thalle ; émettre, sans se marier, des spores germinatives, comme si nous donnions naissance à des enfants à partir de cellules se détachant des orteils ; ou libérer des micro-organismes flottants, mâles ou femelles, différenciés ou morphologiquement ambivalents, qui s’ils sont courtisés, et après un vrai ballet, fusionnent pour créer un œuf.
Des algues vertes plus exposées à la déshydratation, par des techniques de reproduction souvent similaires, sont nées les premières plantes terrestres ; mais même sans déranger les grandes “civilisations végétales” qui ont suivi, les algues ont déjà colonisé les sols, bons ou mauvais, toutes les terres émergées.
Dans les plus petites flaques d’eau, sur les murs, entre les pierres et sur le tronc des arbres, vivent, souvent prises pour des mousses, des milliards d’algues vertes microscopiques. Certaines, comme la Trebouxia et le Nostoc, s’associent de façon permanente avec des champignons pour donner naissance aux lichens, organismes d’assaut, capables de faire face aux froids polaires et aux déserts les plus ardents ; d’autres, portés par les vents, prospèrent carrément dans la neige comme si de rien n’était.
Chlamydomonas nivalis, pour n’en citer qu’un, émerveille chaque année les skieurs avec le phénomène des “neiges rouges”, coloré par le pigment qu’il élabore pour se défendre contre les ultraviolets ; et sur l’Himalaya, à plus de 5000 m d’altitude, c’est encore une algue unicellulaire qui détient, pour le Livre des records Guinness, le record absolu de la vie végétale en altitude.
Les végétaux à la conquête des terres émergées.



Texte © Dr. Giuseppe Mazza

Traduction en français par Michel Olivié
Quelle fut la première plante terrestre ?
Qui peut le dire ? S’il ne fait aucun doute, comme nous l’avons exposé dans le chapitre sur les algues, que la vie sur Terre est née en une seule fois et que donc tous les êtres vivants ont un ancêtre commun, il est plus difficile de reconstituer la grande épopée des végétaux partis à la conquête des terres émergées.
Certes les protagonistes ont été les ancêtres des mousses, des fougères, des lycopodes, des sélaginelles et des prêles, des plantes qui sont toutes plus ou moins étroitement apparentées au monde des algues mais les liens et les origines de ces groupes se perdent dans la nuit des temps.
Quand la nature crée du neuf, quand pour coloniser un milieu elle invente des “prototypes” elle le fait, comme nous, en un petit nombre d’exemplaires et pour une durée limitée. Puis, si l’idée se confirme, elle commence la “production en série” et c’est seulement quand la “recette pour la vie” remporte partout des succès que nous trouvons des fossiles.
Mais les héros du moment, les innovateurs, disparaissent en général sans laisser de traces. C’est comme si dans quelques millions d’années un paléontologue spécialiste en automobiles cherchait dans les roches les vestiges de notre civilisation : il trouvera peut-être, à supposer qu’il se soit fossilisé, le capot de la “coccinelle Volkswagen” mais sûrement pas les squelettes des premières voitures ressemblant à des fiacres et produites artisanalement en nombre limité. Il est donc difficile, sinon impossible, de dire quelle algue verte pluricellulaire a tenté pour la première fois de vivre hors de l’eau, peut-être sur un rocher éclaboussé par les flots ou peut-être au bord d’une paisible lagune.
La “palme des vainqueurs” fut dans un premier temps décernée aux mousses déjà présentes parmi les fossiles du Carbonifère mais probablement plus anciennes compte tenu de leurs dimensions modestes et de leur aspect très semblable à celui des algues du bord de mer.
Et de fait les botanistes du XIXe siècle ont mis triomphalement en évidence la présence chez l’Hépatique (Marchantia polymorpha) d’une “mousse anormale” en forme d’algue qui serait le chaînon assurant la connexion entre cette dernière et les premières plantes terrestres. Mais, par la suite, à partir de la stratégie de reproduction des hépatiques ils se sont rendus compte qu’il s’agissait seulement de mousses régressives, nostalgiques du passé, qui, étant amenées à vivre le long de torrents sur des rochers trempés, n’ont su résister à l’envie de se mettre à ramper comme des algues.
L’évolution mélange les cartes et, pour compliquer les hypothèses, on n’avance jamais en ligne droite mais en zigzag ou comme dans certaines danses populaires où l’on fait trois pas en avant et deux en arrière. Aujourd’hui beaucoup de certitudes se sont écroulées et il y en a même qui soutiennent que les mousses dérivent de plantes palustres inconnues qui ont été miniaturisées par des événements climatiques.
Il vaut mieux, cela étant, rester objectif et s’en tenir aux faits. Regardons de près un tapis de mousse. Même si sous les tropiques il existe de nombreuses espèces retombantes insolites dotées de petites tiges longues jusqu’à un mètre et même si les bryophytes qui regroupent les mousses, les hépatiques et les sphaignes comptent largement 23.000 espèces et donc 23.000 “modèles” différents, il s’agit presque toujours de petites plantes fragiles de quelques millimètres. Elles vivent au coude à coude, appuyées l’une contre l’autre, comme les polypes des coraux de récif, parce que l’union fait la force et qu’avec leur structure en coussinet elles retiennent mieux l’humidité et la quantité d’humus qui suffit pour coloniser même les rochers les plus inhospitaliers.
Observons un seul élément de la colonie. En quoi se distingue-t-il d’une algue ?
Tout d’abord il pousse hors de l’eau. Il a une petite racine qui pompe l’humidité du sol et les liquides passent seulement par osmose d’une cellule à l’autre. Les faire circuler sans tissus conducteurs n’est pas simple et cela explique tout de suite ses dimensions modestes mais ce qui frappe le plus chez les mousses est leur structure d’ensemble en “arborescence” et leurs feuilles qui sont totalement inconnues dans l’univers des algues.
Cette innovation est essentielle au point que les botanistes divisent aujourd’hui le règne végétal en deux sous-règnes : celui des Thallophytes, les “plantes archaïques”, comme les bactéries, les algues, les champignons ou les lichens qui ont un corps comportant une lamelle foliaire plus ou moins différenciée et celui des Cormophytes, les “plantes modernes”, qui possèdent des racines, des tiges et des feuilles disposés savamment dans un espace tridimensionnel.
C’est une invention révolutionnaire si l’on songe que pour effectuer la même photosynthèse un chêne de 25 m de haut devrait avec la méthode des algues exposer au soleil et au vent une énorme lamelle foliaire de 1.200 m².
Certaines petites mousses ont à leur sommet un pédoncule long et fin comportant une capsule. Elle contient les spores. Celles-ci, disséminées une fois mûres par les contorsions du pédoncule qui réagit comme un hygromètre à l’humidité atmosphérique, sont formées d’une seule cellule qui contient la moitié des chromosomes de la plante-mère.
Dans des conditions particulières les spores germent en générant un thalle allongé ressemblant à une algue filamenteuse, le Protonema, d’où sortent les minuscules “arborescences” du tapis qui au printemps portent les organes reproducteurs mâles, les Antéridies, de petites “massues” d’où naissent les spermatozoïdes ou d’étranges “bouteilles” au col long et étroit, les Archégones, qui contiennent la cellule-œuf.
L’initiative, comme toujours, incombe aux mâles. Après la pluie, quand le tapis est bien trempé, ils sortent par milliers en nageant grâce à leurs cils mobiles et rejoignent leurs compagnes afin de les féconder.
Grâce à cette union le patrimoine chromosomique double et l’œuf germe sur place comme un parasite de la plante-mère. Un être sans feuille parce que la mousse dépourvue de vaisseaux ne peut pas l’alimenter en eau. Un petit E.T. avec une seule idée en tête : produire le pédoncule et l’urne avec les spores. Cette urne, dont la forme est particulière pour chaque espèce, présente de minuscules “dents” en bois, hygroscopiques, conçues exprès pour l’ouvrir, à maturité, dans des conditions atmosphérique optimales.
Une stratégie gagnante qui ont permis aux mousses de parvenir presque inchangées jusqu’à nous avec la restriction de la taille et le fait qu’elles ne peuvent se reproduire que quand il pleut.
Leur grand mérite est d’avoir inventé les feuilles et découvert le bois pour les petites dents des capsules mais elles n’ont pas su en tirer parti pleinement comme le vaste monde des fougères.
Celles-ci qui prospéraient à peu près à la même époque avec un maximum de diffusion il y a environ 300 millions d’années ont utilisé en effet la lignine pour construire des organes de soutien et un vrai système circulatoire : des tubes qui conduisent l’eau depuis le sol jusqu’aux feuilles et alimentent en sève sucrée toutes les différentes parties de la plante.
Bien que l’invention des vaisseaux ligneux ait peut-être été due à d’autres groupes aujourd’hui éteints, à une espèce comme l’Aldonophyton antiquissimum découvert seulement en 1953 en Sibérie dans des roches du Cambrien vieilles de 500 millions d’années ou à des plantes sans feuilles comme les Psilophyton et les Rhynia ressemblant à des roseaux et répandus il y a environ 400 millions d’années, c’est principalement avec les fougères que les arbres de la préhistoire ont atteint des performances comparables à celles d’aujourd’hui.
De vraies racines, de vrais troncs et de grandes feuilles. Une association efficace, très souple, qui compte aujourd’hui encore à côté d’espèces géantes, de mini-fougères semblables à des brins d’herbe, des espèces flottant à la manière de radeaux et d’étranges plantes épiphytes, les Platycerium, qui forment sur les arbres tropicaux et dans les serres des fleuristes d’élégants “nids de feuilles” en forme de bois de cerf.
Elles ont toutes en commun la même stratégie de reproduction.
Là aussi, comme pour les mousses, on part de minuscules spores avec un nombre de chromosomes réduit de moitié qui germent en donnant naissance à une lamelle cordiforme appelée Prothalle. Une structure qui ne dépasse pas 3 à 4 cm même chez les espèces de grande taille comme les Dicksiona et les autres fougères arborescentes géantes qui, ressemblant à des palmiers, dépassent 20 m de haut.
Mais à la différence du thalle des mousses cette lamelle ne génère pas d'”arborescence” : elle porte directement en haut les Archégones et en bas les Anthéridies.
Même nage des spermatozoïdes grâce à la pluie mais la nouvelle plante issue du mariage ne vit pas “à l’économie” comme un parasite sur le prothalle, plante ses racines dans le sol et pousse de façon autonome, en abondance, en créant les belles frondes ciselées admirées de tous.
Selon les botanistes ce sont des “feuilles incomplètes” qui en fait fusionnent en une lamelle compacte chez des espèces comme les Asplenium des tropiques ou notre Phyllitis scolopendrium. Jeunes, toutes s’enroulent de façon élégante comme des “houlettes de berger” et portent en général à maturité sur le limbe inférieur des taches foncées, des arabesques formées de nombreux petits points appelés Sporanges. Au microscope ils ressemblent à de petits coffres et s’ouvrent d’une façon similaire aux capsules des mousses en libérant les spores.
Des plantes qui se tournent en arrière vers l’univers des algues, et qui, comme elles, ont besoin d’eau pour se reproduire mais avec des structures déjà modernes et des organes sans cesse plus fonctionnels et spécialisés.
Les prêles qui sont de très proches parents des fougères également connus sous les noms de “queue de cheval”, de “queue de renard” ou de “queue de rat” et à l’honneur entre 200 et 130 millions d’années avec des espèces géantes poussent encore plus loin la”répartition des tâches”.
Prenons le très commun prêle des champs (Equisetum arvense) : le même pied, la même plante possède au printemps des rameaux sans chlorophylle qui portent une sorte de “pomme de pin” allongée remplie de spores et en été des rameaux stériles d’une belle couleur verte qui réalisent la photosynthèse et emmagasinent les sucres dans un organe de réserve souterrain.
Mais les surprises ne s’arrêtent pas là : les spores, parfaitement identiques et dotées de quatre curieux tentacules hygroscopiques appelés Élatères, peuvent donner naissance suivant le terrain où elles tombent à un prothalle mâle ou à un prothalle femelle.
Leur sexe, indéterminé à la naissance, dépend en effet de la richesse du sol : quand il est fertile et donc en mesure de nourrir de nombreuses plantes il naît davantage de femelles et s’il est pauvre les prothalles sont presque tous des mâles qui espèrent qu’une pluie compatissante mène un prince charmant parti à l’aventure jusqu’à la belle qui attend à l’écart dans un coin plus propice.
Au sujet de la fonction des Élatères plusieurs hypothèses ont vu le jour. D’un côté elles servent, comme les voiles face au vent, à aller plus loin et d’autre part en s’entrecroisant en formant une chaîne elles font en sorte que ces étranges prothalles unisexués ne soient pas trop éloignés pour les mariages.
La Selaginella, une plante exotique apparentée aux lycopodes que l’on trouve souvent à cause de ses petites frondes décoratives dans les serres des fleuristes, pousse encore plus loin la différenciation sexuelle en produisant sur un cône, comme les prêles, des milliers de Microspores mâles en haut et 4 grosses Macrospores femelles dotée de réserves de nourriture en bas.
On pourrait donc établir la lignée évolutive suivante : d’abord les fougères aux sexes réunis sur un seul prothalle puis les prêles aux spores non différenciées mais aux prothalles sexués et enfin les sélaginelles aux spores agencées selon leur fonction et aux prothalles sexués.
C’est une belle hypothèse qui va dans le sens d’une efficacité toujours plus grande de la nature si ce n’est que les sélaginelles et les lycopodes répandus aujourd’hui ici et là avec une préférence pour les forêts pluviales denses ont des fossiles plus anciens que les fougères et que leur corps, d’autre part , présente des signes évidents d’archaïsme comme leurs feuilles minuscules et la division dichotomique de leurs branches.
On doit donc en conclure que l’évolution sexuelle des premières plantes terrestres ne s’est pas faite au même rythme que leur apparence.
Certains groupes ont privilégié le sexe, d’autres le port. Mais à l’intérieur de chaque groupe les organes sexuels se sont toutes les fois perfectionnés dans le sens indiqué.
Les ancêtres des sélaginelles actuelles qui se sont répandus avant les mousses et les fougères du Dévonien il y a environ 400 millions d’années n’avaient pas par exemple comme celles-ci des spores et des prothalles aux sexes séparés et certaines fougères aquatiques actuelles comme la Salvinia, bien qu’appartenant à un groupe sexuellement en retard, ont des spores et des prothalles sexués.
Il faut enfin ajouter que si l’évolution récompense en général les précurseurs par une descendance infinie digne des fils d’Abraham les plantes préhistoriques parvenues miraculeusement jusqu’à nous ne sont pas nécessairement les meilleures. Un très important groupe de lycopodes et de fougères, aujourd’hui éteints, a fait par exemple de nouvelles avancées dans la question de la protection de la descendance grâce à l’invention de l’ovule et de la graine et cette nouvelle aventure fera l’objet du prochain article.
C’est ainsi que les plantes ont découvert l’amour.



Texte © Dr. Giuseppe Mazza
Traduction en français par Michel Olivié
Un des grands axes évolutifs de la nature est la protection de l’enfance. Après avoir débarqué sur la terre ferme et inventé les feuilles et le bois il y a environ 300 millions d’années les plantes ont fait un saut qualitatif dans cette direction.
Il ressort de l’examen des fossiles qu’un parent très proche de la Selaginella, le Lepidocarpon, une espèce éphémère qui s’est éteinte au Carbonifère, ne dispersait plus sur le sol ses 4 macrospores. Trois avortaient et la quatrième germait sur la plante en donnant naissance à un prothalle protégé par le macrosporange et une écaille.
Parallèlement, à la même époque, à côté des fougères normales apparurent d’autres fougères, a première vue identiques, qui accentuèrent cette tendance. Le macrosporange y est enveloppé de plusieurs petites feuilles qui se soudent pour former un “tégument” et se transforme lui-même en un organe de protection appelé “nucelle” (du latin “petite noix”) à l’intérieur duquel le prothalle femelle se développe. Une structure nouvelle et compacte qui offre une double protection au futur nouveau-né et qui est appelée “ovule”.
Les microspores germent aussi sur la plante en donnant naissance à des grains de “pollen”. Le prince charmant n’arrive plus à la nage mais avec ces mini-capsules spatiales transportées par le vent.
Il atterrit sur un minuscule orifice de l’ovule appelé “micropyle” (du grec “petite porte”), pénètre dans le nucelle et rejoint la belle pour la féconder. Le mariage s’effectue ainsi loin du milieu extérieur, à l’abri de la sécheresse qui sévit de façon extrême au Permien et alors que les étangs et les lacs s’assèchent du fait de la diminution des précipitations.
Les “fougères à ovules” se sont éteintes vers la fin de l’ère secondaire et pour avoir une idée de leurs grands organes reproducteurs nous devons aujourd’hui nous tourner vers une plante qui vivait à la même époque, le Ginkgo (Gingko biloba) qui a le rare privilège d’être le seul représentant d’une Classe, d’un Ordre, d’une Famille et d’un Genre. Un “fossile vivant” parvenu miraculeusement intact jusqu’à nous depuis le Jurassique.
Ses belles feuilles en éventail se divisent en deux lobes, signe incontestable d’archaïsme (la structure dichotomique, comme nous l’avons vu, date des algues) mais le fait qu’elles tombent en automne après avoir recouvert l’arbre d’une cascade de pièces d’or est déjà un caractère des temps modernes. Les sexes sont séparés et, cas unique dans le monde végétal, tout de suite reconnaissables à leur aspect. Les ginkgos mâles sont en effet élancés tels des sapins alors que les femelles sont plus ramifiées et plus basses.
Mr Ginkgo, au printemps, confie au vent un nombre incroyable de grains de pollen tandis que Mme Ginkgo les attend avec une gouttelette de liquide visqueux sur ses micropyles. Les grains de pollen y adhérent et elle recueille alors le liquide dans une petite chambre nuptiale où les sphères volantes s’ouvrent. De chacune sortent deux spermatozoïdes mobiles qui mettent six mois pour rejoindre l’archégonium. Entre-temps l’ovule, une sorte de cerise jaune, sera tombée au sol.
À peine fécondé l’embryon grandit aussitôt et à foison en utilisant les réserves de l’ovule et c’est là la grande différence avec les plantes modernes à graines, les graines peuvent attendre pour germer jusqu’à des centaines d’années mais non les ovules. Une différence subtile qui jusqu’au début du siècle passé n’avait pas été perçue par les botanistes.
L’invention de l’ovule et celle de la graine sont deux étapes séparées d’environ 30 millions d’années.
Si l’on veut établir une comparaison avec le monde de la zoologie les plantes à ovules sont l’équivalent des animaux ovipares alors que les plantes à graines, plus évoluées, le sont des animaux placentaires. La graine reste en effet liée jusqu’ à sa maturité à la plante-mère où elle puise sa nourriture grâce à un organe que les botanistes appellent, non par hasard, “placenta”. Mais surtout la graine, avec son embryon à la vie latente, permet aux plantes cette “fuite dans le temps” qu’est le sommeil et pour l’instant inaccessible à l’homme: pouvoir dormir pendant des siècles, sans vieillir, au cours de longs voyages spatiaux ou dans l’attente que les progrès de la médecine trouvent un remède à des maladies aujourd’hui incurables.
Certaines graines prélevées dans des herbiers vieux de plus de trois siècles se sont avérées fertiles et d’autres, conservées au frais dans des tourbières, ont germé 1.000 ans après. Un avantage supplémentaire des plantes à graines réside dans le fait qu’elles portent à maturité seulement des œufs féconds et qu’elles ne gaspillent donc pas d’énergie, comme les poules, à fabriquer des œufs stériles.
De ces “plantes ovipares” autrefois très répandues nous avons encore quelques représentantes dans la famille des Cicadaceae. Ici aussi les deux sexes sont séparés.
Mme Cycas revoluta dont la parenté avec les fougères arborescentes à ovules paraît probable (aspect analogue et feuilles qui croissent enroulées à l’apex) met elle aussi en oeuvre la technique de la gouttelette visqueuse. Ici les ovules ne sont pas portées par un pétiole mais se développent sous des feuilles spéciales de couleur dorée repliées sur elle-mêmes en formant une sorte de chou. Pendant dix jours quand Mme est féconde ces feuilles se soulèvent légèrement pour laisser entrer le pollen et ensuite elles se referment en portant même à maturité, comme chez le Ginkgo, les ovules qui n’ont pas eu la chance de le rencontrer.
Encore une fois, comme dans les contes, la vie naît dans les choux et dans cette étrange structure protectrice foliaire ni molle ni dure les botanistes devinent déjà le “projet” assigné aux pommes de pin des conifères qui se borneront à miniaturiser cette ossature tout en la rendant plus solide.
En fait même le sexe de Mr Cycas revoluta n’est pas très différent de celui d’une pomme de pin. Il se dresse sous la forme d’une structure fusiforme et pointue de 30 à 40 cm et brûle d’un amour tel que sa température interne s’élève de 10 °C. Il soulève ses écailles, expose les sacs polliniques et libère un peu à la fois dans le vent 5 milliards de grains de pollen.
Visibles à l’œil nu les spermatozoïdes ciliés de cette espèce sont les plus grands de la nature : ils mesurent environ un tiers de millimètre et mettent 4 mois pour rejoindre la cellule femelle à l’intérieur de l’ovule. Ici aussi la fécondation proprement dite s’effectue ensuite souvent sur le sol.
Les Cicadaceae, qui étaient répandues au Mésozoïque au point de représenter plus d’un tiers de la flore terrestre, sont aujourd’hui dispersées dans les zones tropicales et subtropicales et comptent dix genres (une onzième semble avoir été découverte en Colombie) et environ 130 espèces. Ce sont toutes des plantes dioïques qui confient leur pollen au vent mais certaines espèces sud-africaines comme l’Encephalartos villosus et l’Encephalartos altensteinii ont passé aussi un traité avec les insectes pour le transport du pollen.
Leur partenaire est un curieux coléoptère curculionide, l’Anthiarhinus zamiae. Ses femelles visitent les cônes mâles, attirées, semble-t-il, par la chaleur et l’odeur qu’ils dégagent, après quoi, enfarinées comme il se doit, elles vont sur les cônes femelles pour y pondre leurs oeufs. Elles explorent chaque fente du strobile en fécondant les ovules avec leur longue trompe et même si leurs larves en détruisent quelques-unes, sur le plan de l’évolution, cela reste un grand pas en avant par rapport à la pollinisation anémophile. C’est peut-être ainsi qu’a commencé il y a des millions d’années la captivante collaboration entre les insectes et les plantes.
La croissance des Cicadaceae, reléguées pour la plupart dans des lieux inhospitaliers où la vie est difficile et où la concurrence avec les plantes modernes est donc moindre, est incroyablement lente, jusqu’à 5 à 10 cm par an pour les espèces peu nombreuses qui survivent dans les climats chauds et humides et moins de 0,5 cm pour celles des milieux arides. Si l’on ajoute à cela que la pollinisation est souvent difficile parce que les plantes des deux sexes sont trop éloignées les unes des autres on peut comprendre aisément pourquoi depuis des temps anciens les activités humaines ont donné le coup de grâce au reliquat de ces plantes préhistoriques.
Les graines de la Macrozamia spiralis ont été systématiquement récoltées par les aborigènes australiens pour en extraire une farine et les malheureux Encephalartos d’Afrique du Sud n’ont assurément pas connu un meilleur sort vu que les indigènes, en plus de leurs graines, consommaient la partie supérieure des tiges riche en amidon (Encephalartos vient de EN = à l’intérieur, KEPHALE = tête et ARTOS = pain).
À la différence du Ginkgo protégé depuis l’Antiquité par l’homme qui le plantait en en faisant des bosquets autour des temples bouddhistes et le vénérait, convaincu qu’il écartait les risques d’incendie, les Cicadaceae, qui sont pour la plupart originaires de milieux pauvres, ont été presque partout détruites brutalement.
Une rare exception existe en Afrique du Sud, dans l’ancien Lebowa, avec la spectaculaire forêt d’Encephalartos transvenosus de Modjadji, à environ 300 km au Nord de Johannesburg. Là, sur une montagne sacrée placée depuis des siècles sous la protection de la Reine des pluies, on trouve la plus grande concentration de Cicadaceaeau monde. Certains arbres hauts de 12 à 13 m ont plus de 1.000 ans. Les sentiers soigneusement agencés de la réserve actuelle ne portent en rien ombrage à la fascination d’un plongeon dans le Mésozoïque.
Une autre grande espèce du Natal, l’Encephalartos woodii, par contre, est aujourd’hui malheureusement éteinte dans la nature. On n’ a sauvé que deux mâles adultes qui sont placés sous une étroite surveillance dans le jardin botanique de Durban et même si de quelques bourgeons en dormance on a isolé deux ou trois petits plans, comme il n’y a plus de femelles on est en droit de dire que c’est l’espèce d’arbre la plus rare au monde.
Mais tournons-nous vers l’évolution et l’invention de la graine qui est, semble-t-il, l’oeuvre de plantes apparentées aux conifères.
Prenons par exemple l’If. C’est une espèce dioïque comme le Ginkgo et les Cicadaceaemais à la différence de ceux-ci elle a un grain de pollen plus évolué.
Après avoir rejoint le micropyle de l’ovule il ne produit plus de spermatozoïdes ciliés mais ceux-ci traversent le nucelle, plus ferme, au moyen d’un “tube pollinique”. Après la fécondation l’embryon cesse jusqu’à un certain point de se développer et se déshydrate en attendant qu’arrive le meilleur moment pour germer. Entre- temps son tégument se transforme sous la forme d’une excroissance charnue, l’arille, qui le fait ressembler à une baie rouge, d’où le nom scientifique de l’espèce, Taxus baccata.
Ce n’est pas encore un fruit mais il en est très proche. Les oiseaux en sont friands et le transportent loin au grand soulagement de Maman If qui ne risque pas ainsi d’entrer en concurrence avec ses fils.
Les Pins et les Mélèzes, qui sont plus modernes par certains aspects, portent les deux sexes sur la même plante (il est curieux qu’ à l’origine les sexes étaient séparés sur les plantes et réunis chez les mammifères et que par la suite l’évolution a inversé cet état de choses). Les microsporanges et les macrosporanges ont été rassemblés dans les structures coniques que l’on connaît tous sous le nom de pommes de pin.
Les cônes mâles émettent des nuages de pollen confiés au vent alors que les cônes femelles ont deux ovules par écaille. Celles-ci, comme chez les Cicadaceae, se soulèvent pour la pollinisation et se referment ensuite en se soudant les unes les autres dans une structure compacte appelée en général à s’ouvrir la seconde année afin de disperser les graines.
Une formule gagnante qui a engendré tous les conifères que nous connaissons et qui sont répandus dans des régions très froides mais aussi sous les tropiques comme les Araucarias qui forment des forêts denses dans l’hémisphère Sud.
L’étape suivante, la protection de la graine dans un fruit, se laisse déjà entrevoir chez deux autres gymnospermes ( du grec “graine nue”), la Welwitschia et l’Ephedra, des plantes très différentes et dernières représentantes des groupes qui ont assuré la transition vers les Angiospermes (du grec “graine à involucre”). Chez la Welwitschia qui a survécu par miracle dans le désert de Namibie (voir le reportage publié dans SCIENZA § VITA) les ovules sont déjà enveloppés par un “sac protecteur” qui se resserre en partie haute sous la forme d’un tube qui reste ouvert afin que le pollen puisse rejoindre le micropyle.
Chez les Ephedra, de petits arbustes ligneux ramassés en Chine depuis des millénaires pour en extraire un alcaloïde, l’éphédrine, aux propriétés toniques et anti-asthmatiques, les ovules, dotés d’un bec, sont enfermés dans une “bouteille” au long col qui s’épaissit après la fécondation en formant un fruit charnu rouge ou jaune.
On se dirige vers des plantes dont l’ovule, totalement séparé du monde extérieur, est fécondé à travers un style et un long tube pollinique. Des plantes davantage liées au monde animal et qui pour la plupart réuniront les deux sexes dans une structure unique, la “fleur”, dont nous parlerons dans le prochain article.
Quand les végétaux ont inventé les fleurs.



Texte © Dr. Giuseppe Mazza
Traduction en français par Michel Olivié
À chaque grande découverte du monde des végétaux correspond la naissance d’une civilisation végétale.
Grâce au bois les fougères dominèrent la flore du Carbonifère et du Permien, entre 350 et 220 millions d’années, puis, à l’ère secondaire, elles furent supplantées par les premières plantes à graines, les Gymnospermes, qui célébrèrent leur triomphe au Jurassique, il y a 150 millions d’années, avec environ 20.000 espèces de conifères. Aujourd’hui elles se sont réduites à moins de 600 espèces et les fougères qui ont proportionnellement mieux résisté à l’assaut du temps doivent pour la plupart se contenter de tailles modestes.
Les Thallophytes mises à part, neuf espèces actuelles sur dix appartiennent à un nouveau genre de plantes, les Angiospermes, nées dans les forêts tropicales au début du Crétacé il y a 130 millions d’années.
Ces plantes très compétitives et agressives peuplèrent rapidement les tropiques et se propagèrent ensuite par manque d’espace vers les régions tempérées. En 20 millions d’années elles atteignirent l’Europe et 30 millions d’années plus tard elles envahirent avec plus de 250.000 espèces toutes les terres émergées.
Quelle fut leur carte maîtresse?
Tout d’abord l’invention de l’ovaire, une “maison” d’une ou plusieurs pièces où les ovules sont protégées et vouée à se transformer en fruit après la fécondation et en second lieu l’alliance avec les animaux, spécialement les insectes, pour le transport du pollen.
Pour les séduire ces plantes se sont en un certain sens “animalisées” en créant une structure multicolore, la fleur, tout-à-fait étrangère à leur monde végétal.
Des feuilles qui se colorent vers l’extrémité des rameaux, comme on peut l’observer chez des espèces encore “indécises” comme l’Étoile de Noël, dans le but d’assumer une fonction différente de la photosynthèse, des pétales qui se transforment progressivement en étamines comme chez les Nénuphars où en se déplaçant vers le centre de la fleur ils s’amincissent jusqu’à devenir les pédoncules des anthères.
L’architecture de la corolle découvre au premier regard l’animal à séduire :des “Fleurs à oiseaux” de grandes dimensions, pour la plupart rouges ou violettes comme les Passiflora et les Hibiscus qui, riches en nectar, portent leurs organes sexuels sur un pédoncule, loin du bec de leurs partenaires affamés, ou comme la Freycinetia funicularis qui, pour se reproduire, offre stoïquement aux oiseaux une salade de bractées charnues et succulentes, des “Fleurs à souris”, riches en sucre, comme la Banksia petiolaris et d’autres Protéacées qui fleurissent au ras du sol, des “Fleurs à chauves-souris” comme celle du Baobab ou de l’inquiétant Couroupita guianensis, l’Arbre aux boulets de canon aux corolles coriaces et aux organes reproducteurs sensuels et animaux ressemblant aux tentacules d’une Anémone de mer, et naturellement les “Fleurs à insectes” comme la Marguerite ou la Sauge qui sont souvent dotées de systèmes de pollinisation sophistiqués.
Il y a des plantes cruelles comme certaines orchidées qui torturent et trompent les guêpes, des plantes comme certaines Aracées qui emprisonnent et tuent les malheureux “facteurs du pollen”, mais aussi des plantes gentilles comme le Marronnier (Aesculus hippocastanum) qui, pour faciliter la tâche des abeilles, signalent la présence du nectar dans ses fleurs au moyen d’une petite tache jaune qui vire au rouge quand le “réservoir” est vide.
Mais la Nature ne renonce jamais totalement aux anciennes méthodes employées avec succès pendant des milliers d’années et, à côté des Angiospermes zoophiles il existe aussi beaucoup d’Angiospermes anémophiles “conservatrices” qui, vivant côte à côte dans de vastes rassemblements où la pollinisation est aisée ou dans des climats froids où les insectes se font plus rares, continuent, comme la société des Conifères qui les a précédées, à utiliser le vent pour le transport du pollen.
C’est le cas des Graminées, le Maïs (Zea Mays), par exemple, aux étamines portées par de longs filaments qui frémissent au moindre souffle de vent, et des “plantes à chatons” comme le Noisetier (Corylus avellana) ou le Peuplier (Populus nigra) aux fleurs réunies en épis ondoyants qui sortent avant les feuilles pour mieux disperser, sans nul obstacle, des quantités impressionnantes de pollen. 5 millions de grains pour un seul chaton de Bouleau, 500 millions par saison avec la conséquence bien connue des allergies pour un seul pied de Noisetier.
Dans ces cas les corolles bien évidemment ne servent pas puisqu’il n’y a aucun partenaire à séduire mais il s’agit au final de minorités très faibles.
Pour les Angiospermes types les pétales qui sont l’enseigne publicitaire de la fleur placée autour des organes sexuels font de la réclame pour son bon nectar suivant un schéma bien précis. À l’extérieur des feuilles transformées, les sépales qui protègent les fleurs en bouton, au centre l’ovaire qui contient les ovules hérités du monde des Conifères et protégés par plusieurs enveloppes inventées par la société des fougères et à l’intérieur la précieuse cellule femelle héritée de la société des algues. Dans l’appareil reproducteur des plantes à fleurs on peut lire, comme dans les diverses composantes du cerveau des vertébrés supérieurs, l’histoire de l’évolution.
Les organes mâles, appelés étamines, ont deux microsporanges, les anthères. Ce sont les “testicules” des plantes qui génèrent des milliers de grains de pollen, des “mini-navettes spatiales” conçues pour atterrir sur le “stigmate”, la partie terminale du “style”, une sorte d’antenne de télévision directionnelle ou parabolique reliée à l’ovaire où se trouve la cellule femelle.
Éveillé par une sécrétion hormonale le pollen germe et émet pour la rejoindre un long “tube” qui se fraie un chemin dans le style. Un procédé nouveau qui était déjà présent chez les Conifères mais, en comparaison, les Angiospermes sont beaucoup vaillants et les mâles avancent 1.500 à 2.000 fois plus vite.
Une fois l’ovule atteint deux spermatozoïdes sortent du tube et il se produit une chose très étrange, caractéristique de ces plantes et tellement particulière qu’elle fait penser que toutes les Angiospermes ont eu un ancêtre commun. Un des deux spermatozoïdes s’unit à la cellule femelle pour créer comme d’habitude un oeuf avec 2n chromosomes alors que l’autre fusionne avec deux cellules du sac embryonnaire pour créer un oeuf spécial à 2 n chromosomes. Les deux entités évoluent séparément mais alors que le premier oeuf génère l’embryon le second, nourri à travers le placenta par la plante-mère, forme un tissu à trois éléments chromosomiques, appelé “albumen”, un véritable lait concentré très énergétique destiné à nourrir la jeune plante au commencement et à lui donner la force de se constituer une racine.
Ainsi le père, bien qu’à titre posthume, contribue, à la grande différence de ce qui se passe dans le monde animal, à l’alimentation de sa descendance.
Pour le reste les aventures amoureuses du monde végétal ne diffèrent guère des nôtres. Aux mâles incombent l’initiative, l’aventure d’un voyage difficile et une rude compétition avec des rivaux pour la conquête de l’épouse alors que les femelles, même si elles restent “à la maison” dans le ventre de la fleur, tirent les ficelles et comme toujours maîtrisent habilement la partie.
Les papilles de leurs stigmates écartent d’abord mécaniquement, comme une serrure le ferait pour de fausses clés, les graines des autres espèces et émettent ensuite des sécrétions sucrées spéciales excitantes uniquement pour leur type de pollen. Mais elles ne s’arrêtent pas là. Quand débute la grande compétition des tubes polliniques elles livrent une guerre chimique aux consanguins et une fois qu’elles ont choisi le prince charmant, celui dont le contenu chromosomique est le meilleur, elles l’aident dans son parcours par des émissions hormonales particulières.
Bien sûr, avec les deux sexes réunis dans une structure unique, l’inceste est toujours aux aguets mais les avantages son énormes. En effet, quand ils sont séparés, ce qui arrive seulement dans 10 % des cas avec des espèces dites “dioïques” comme les Saules, les Peupliers ou l’Ortie les plantes mâles, après la fécondation, ne servent plus à rien et sur un emplacement équivalent la production de graines est réduite de moitié.
Il vaut donc mieux prendre le risque de choisir, même si des handicaps peuvent en théorie s’y ajouter, des caractères qui sont favorables d’autant plus que la Nature élimine ensuite promptement les produits défectueux en effaçant ses erreurs par la mort.
Il y a beaucoup d’espèces comme la Nigella damascena où si l’époux ne parvient pas à un certain point le stigmate se replie vers les anthères pour s’auto-féconder.
Les plantes à fleurs s’efforcent en général par diverses techniques de réduire le plus possible les accouplements consanguins. Certaines espèces comme le Thym, la Lavande et certaines variétés de Pommes et de Poires sont auto-stériles : c’est-à-dire qu’elles reconnaissent par une substance protéique leur propre pollen et les empêchent de germer. Une autre stratagème consiste à faire mûrir les étamines et les pistils à des périodes différentes de telle sorte que les femelles ne soient réceptives que quand leurs frères ont disséminé tout leur pollen. Chez les fleurs à insectes il existe souvent des “séparations mécaniques” très efficaces.
Les Angiospermes les plus anciennes en se basant sur les fossiles n’appartiennent pas à une lignée commune mais à des groupes très différents comme les Palmiers aux inflorescences souvent énormes, les Saules aux fleurs réduites à leur plus simple expression (anthères ou styles) et adossées à un chaton rigide à la manière des pommes de pin mais déjà dotées d’un nectar destiné à attirer les insectes et les Magnolias qui, avec une disposition similaire dans le sens vertical, portent au centre sur une structure conique les styles entourés en partie basse par les étamines. Celles-ci tombent ensuite et les fruits rouges proéminents viennent au monde sans protection.
Le réceptacle voyant des Magnolias est également présent à une échelle réduite chez les Renoncules et les Fraises où les “pépins” sont en réalité les fruits. Celui que nous mangeons est leur support mais il tend à se réduire chez des plantes plus évoluées jusqu’à devenir plat chez la Reine des Prés et concave avec un petit orifice chez les Roses pour finalement se replier sur lui-même chez le fruit du Pommier qui offre ainsi un maximum de protection aux graines.
Les fruits des Angiospermes sont extrêmement variables et même si chaque famille de plantes a en général des idées précises sur ce point il n’existe pas une étroite corrélation avec la forme de la fleur.
Les botanistes distinguent tout d’abord les fruits secs des fruits charnus.
Parmi les premiers ils séparent ensuite ceux qui sont durs et compacts, appelés “akènes”, comme le Tournesol ou le Pissenlit, de ceux qui sont déhiscents comme les “Capsules” des Iris, des Pittosporums, des Coquelicots, les “gousses” des petits Pois, les “Siliques” de la Giroflée et les “Follicules” du Laurier rose.
Parmi les seconds ils distinguent les “Baies”, les fruits aux graines situées à l’intérieur comme le Raisin, la Groseille ou les figues de Barbarie, les “Drupes” qui ont un seul noyau comme les cerises, les abricots ou les noix et le “Piridion”, une forme intermédiaire entre la drupe et la baie, caractéristique de la Pomme et de la Poire.
Si, mis à part le vent, les plantes se servent principalement des insectes pour le transport du pollen c’est aux vertébrés qu’elles confient au moyen de fruits goûteux et aux couleurs souvent attractives le soin de disperser leurs graines sous forme de déjections après un sombre parcours intestinal, de noyaux non comestibles laissés loin à l’écart de la plante-mère, de réserves d’aliments en excès et abandonnées par inadvertance ou par accident.
Il existe, comme nous l’avons vu dans ces chapitres, une étroite corrélation entre tous les êtres vivants nés presque certainement dans la nuit des temps d’un ancêtre commun.
Si l’on cherche à avoir une vision d’ensemble, ce qui est de plus en plus difficile de nos jours où la super-spécialisation conduit les scientifiques à devenir “myopes” jusqu’à “tout savoir de rien” on se rend compte que le “monde vert” de la chlorophylle n’est pas si éloigné de notre “monde rouge” du sang.
Cela nous donne une leçon d’humilité et de fraternité comme celle, par exemple, de St-François d’Assise et on entrevoit un dessein général, une intelligence de la Nature dont la nôtre est un corollaire conscient.
On comprend que la Création ou l’Évolution ne sont pas des faits du passé mais se modifient chaque jours sous nos yeux avec une logique qui n’est pas toujours la nôtre en suivant, comme le fait un ordinateur, un “programme” que nous n’avons pas créé nous-mêmes.
Comment seront les plantes du futur ?
C’est difficile à dire et en l’état actuel des choses cela dépendra aussi de l’homme. Il semble cependant évident que le “fil d’Ariane” de l’histoire des plantes est la protection de l’ovule et donc de leur descendance et que dans les forêts tropicales, qui sont depuis toujours des laboratoires de la vie, poussent peut-être déjà les espèces qui domineront la Terre de demain.
Voulons-nous tenter à ce sujet une anticipation de cent millions d’années ?
Dans l’obscure famille des Monimiacées, de lointaines parentes des Magnolias, l’ Hennecartia de l’Argentine et la Tambourissa de Madagascar ont déjà inventé par exemple une nouvelle protection interne pour les fleurs : un édifice où elles fleurissent en sécurité et où les fruits “préemballés” se développent tranquillement. Mais notre figuier aussi est une plante d’avant-garde. Ses minuscules corolles poussent à l’intérieur d’une structure charnue que nous appelons “fruit” et qui a une petite porte d’entrée destinée au pollinisateur alors que les fruits véritables, bien protégés, sont les “petites graines” que nous trouvons à l’intérieur.
SCIENZA & VITA NATURA – 1992