
Text © DrSc Giuliano Russini – Biologist Zoologist

English translation by Mario Beltramini

The Trimeresurus albolabris is a crotalid without rattle © Dr. Giuseppe Mazza
Class: Reptilians (Reptilia).
Subclass: Lepidosaurian (Lepidosauria).
Order: Squamates (Squamata).
Suborder: Ophidians or Serpents (Ophidia) or (Serpentes).
The class of Reptilians (Reptilia) includes 3 subclasses:
►►►Archosaurians (Archosauria) with one only order, the Crocodiles (Crocodilia), to which belong also the alligators, the caimans and the gavials.
►►►Lepidosaurians (Lepidosauria), which form the order of the Squamates (Squamata), subdivided in the suborders of the Saurians (Sauria) and of the Ophidians or Serpents (Ophidia or Serpentes).
►►►Anapsidians (Anapsida), which form the order of the Chelonians (Chelonia), the turtles.
In this introductory, we shall treat only of the suborder of the Serpentes subdivided in 3 infraorders:
►►Caenophidians (Caenophidia)
►►Henophidians (Henophidia)
►►Scolecophidians (Scolecophidia)
These three infraorders are subdivided in 9 families:
►Typhlopids (Typhlopidae)
►Leptotyphlopids (Leptotyphlopidae)
►Aniliids (Aniilidae)
►Acrochordids (Acrochordidae)
►Boides (Boidae)
►Colubrids (Colubridae)
►Elapids (Elapidae)
►Hydrophiids or Sea snakes (Hydrophiidae)
►Viperids (Viperidae)
To these nine families do correspond 25 subfamilies, which we shall briefly describe along this text, and several genera, species and races or subspecies. Even if comprising all the families and subfamilies, this treatise will be general, and the deepening of the species will take place in the single texts. The last part of this writing will treat of those physiological aspects of organ and general systems and of the ecology-ecophysiology typical of the ophidians or serpents (for instance, the venom apparatus), as we have already briefly treated about the general characteristics for the class of the reptilians when talking about the Chelonia. Finally, we shall do some brief notes, even if I am not, may be, an expert as a herpetological zoologist, about some pathologies affecting the serpents and the reptilians, both in natural conditions and of life in controlled environment (zoological garden and aquatic park), keeping well in mind that this matter is quite recent and few veterinarians are interested about it.
Infraorder of the Scolecophidians (Scolecophidia)

The Typhlops are primitive burrowers. Some do not even consider them serpents © Dr. Giuseppe Mazza
It includes the family of the Typhlopids (Typhlopidae) and that of the Leptotyphlopids (Leptotyphlopidae).
Similar to small, much specialized, burrowing serpents, the typhlopids are spread in all the warm regions of the planet. Their head is short, not distinct from the neck, the body is cylindrical, elongated and thin, and finally, the tail is short. They are called blind snakes by the herpetological biologists due to the very marked atrophy of their eyes, or also worm serpents, even if this name is quite less justified.
These animals have in the same time some primitive characters, and others indicating a marked specialization. Their systematic position has therefore originated many discussions among taxonomical biologists, and they have even reached the position to doubt about their belonging to the suborder of the Serpents (Serpentes).
Unluckily, the paleontological data available to the biologists are of no help, as the most ancient fossils known, of the European Miocene, already belong to the genus Typhlops. The only sure thing is that the typhlopids occupy presently an isolated position.
Their ecological adaptations to the burrowers’ life are of various types, even if they are not so marked like what happens for the members of the family of the Amphisbaenids (Amphisbaenidae), infraorder Amphisbaenians (Amphisbaenia), suborder Saurian (Sauria), and order Squamate (Squamata).
Usually, the muzzle is dorsoventrally flattened, but in some species, for instance in the African species Typhlops coniagui, the rostral scale, prominent and sharp, forms a vertical angle.

When annoyed the Cylindrophis rufus raises the tail as a head, like a cobra © Dr. Giuseppe Mazza
The small inferior jaw, usually toothless, is placed neatly backward.
The body is covered by scales, all identical, very smooth and much imbricate.
The very short tail is conical, the most of the times as long as the width of the base. It ends in a pointed scale, which surely is utilized as support point during the digging.
The epidermal glands, placed at the base of the scales, are particularly developed, and the same thing happens for the cloacal gland, uneven and median, which occupies most of the tail. The hemipenes (paired organ, in number of two), are extremely long and thin. The uniform colour, but often paler on the belly, goes from the olive yellow to black.
The typhlopids do not have the slow and lazy gait of the amphisbaenids, and not even their locomotion technique. On the contrary, they are very lively animals, difficult to catch, especially the smallest species. The biggest member of the family is the Typhlops punctatus of tropical Africa, which reaches the 80 cm, but most of the species have a length of 15-30 cm, and there are some even smaller.
The family of the Typhlopids (Typhlopidae) is subdivided in two subfamilies: the Typhlopins (Typhlopinae) and the Anomalepines (Anomalepinae).
Most of the typhlopins, belonging to the ubiquitous genus Typhlops and to the South American genus Typhlophis, is characterized by the small size of all the cephalic scales.
In northern America, the typhlopins do not reach the USA, whilst in the Old World there is one species, the Typhlops vermicularis, which lives in Greece, Asia Minor and Middle East. Also in South Africa, Japan and Australia, there are Typhlops living in Mediterranean climate regions, whereas it seems that they are not found in Tasmania.

Maybe the ugliest serpent, but the skin of Acrochordus javanicus is much appreciated by the industry © G. Mazza
In any case, the characteristic habitat of the typhlopins is the more or less humid ground, easy to burrow, of the tropical and equatorial forests and where they can abundantly find the ants and termites which form the preferred dish of their diet.
The biggest species nourish also of small invertebrates, forming the “pedofauna”, but their mouth, narrow and little expandable, does not allow them to swallow big preys.
In the zones where the dry season is more evident, the take shelter in the deep ground waiting for the rains, but usually they live in the surface layer and are often found under the logs and the decaying trunks or at the base of the termite mounds.
It really seems that the typhlopins surface regularly during the night, especially after the rain. They move without any difficulty on the ground, ready to rapidly hide by the least alarm.
Like for the amphisbaenids, we ignore the exact moment of these exits, and if they do this only for hunting or for changing of location.
Little is known about these serpents. Perhaps some typhlopins are oviparous, possibly most of them, even if some species such as the Diard’s blind snake (Typhlops diardii) of south-eastern Asia, are viviparous.
The subfamily of the Anomalepines (Anomalepinae), represents a small primitive group, localized in South America.

The Python sebae lives in various habitats south of Sahara, where may be 7 m long © Dr. Giuseppe Mazza
The head is covered by scales more diversified than those of the typhlopins, which recall the cephalic plates of the Colubrids (Colubridae).
Furthermore, some of them, for instance in the genus Anomalepis, have one tooth on each side of the mandible. However, their general look and behaviour are not much different from those of the other members of the family.
The members of the family of Leptotyphlopids (Leptotyphlopidae) have a look so much similar to that of the Typhlopids (Typhlopidae), especially to the genus Typhlops, that even for an experienced herpetological biologists it is practically impossible to distinguish them at naked eye.
However, we may keep in mind that the leptotyphlopids tend to be thinner, with a little less short tail, which is often longer than wider. But it is sure that the two families have a different origin, as shown by the structure of the skull, the localization of the teeth on the mandible, and the absence of teeth in the upper jaw.
Though there is a quite evident specialization for the hypogeous life, for many other characters the leptotyphlopids are more similar to the recent serpents than what the typhlopids are.
No fossil form is known. The genus Leptotyphlops, the only one of the family, is found in the most part of the tropical regions of the world, but the south-eastern Asia and Oceania.

Menaced, the Python regius folds on itself, head in the middle, protected by the tight spires © G. Mazza
It does not go further east of India, and is not found either in Madagascar.
In America, there are two species living in USA: the Leptotyphlops dulcis, in the centre of USA, south of Kansas, and the Leptotyphlops humilis the whole south-west of the country, that is, in more or less dry zones, with rigid winter.
On the contrary, in the Old World, the leptotyphloids do not go further north of northern Sahara, as it happens for instance for the Leptotyphlops macrorhyncus.
This geographical distribution is the contrary to that of the typhlopids, which are absent in USA, but, as seen, are found in Greece and Asia Minor.
The ranges of the two families over pose amply in the inter-tropical American and African regions, where live most of the species. After the little we know, the habits and customs of the leptotyphlopids are the same as those of the typhlopids.
It seems however that the two American species get out by the sunset and do this with regularity.
The biggest leptotyphlopids do not overcome the 30 cm of length, most of them measure 12-20 cm, whilst others hardly reach the 8-10 cm, with a body as thick as a lead pencil.
The colour, uniform, is usually brown or sand, pink at times.
In some species such as Leptotyphlops macrorhyincus, the rostral scale is very prominent, but the muzzle does never form a real “sagittal crest”.
It seems that leptotyphlopids are oviparous.
Infraorder of the Henophids (Henophidia)

The Python molurus bivittatus lives in India, Indonesia and Southeast Asia, where reaches 6 m © Mazza
It includes the families of Aniliids (Aniliidae), of the Acrochordids (Acrochordidae) and of the Boides (Boidae).
The family of the Aniliids (Aniilidae) is subdivided itself in three subfamilies, much different between them, of primitive burrowing serpents more or less similar to the boides.
A certain number of fossil findings of the upper Cretaceous and of the Eocene, in particular some isolated vertebrae, have been classified as belonging to aniliids. A better conserved serpent of the Argentine Upper Cretaceous, 2 m long, and belonging to the genus Dinilysia, presents intermediate characters between the subfamily of the Aniliines (Aniliinae) and the boides.
The aniliines are medium sized serpents, with the conical head not distinguished from the neck, the cylindrical body, quite often somewhat dorsoventrally flattened in the rear part, and a short and conical tail. The eyes are small but functional. The scales of the body, broad and smooth, are all identical, but some species having a ventral row with slightly bigger scales.
Remains of pelvic girdle are present and, on each side of the cloaca, there is prominent a small hook, like in the boides. This subfamily lives in two well distinguished geographic regions: the tropical South America, with the genus Anilius, and the south-eastern Asia and Indonesia with the genera Cylindrophis and Anomalochilus. The Anilius scytale, which reaches the 90 cm of length, is called “false coral snake” due to its bright colouration, with alternatively red and black rings.
The Cylindrophis, are a little smaller (50-70 cm) brown or black, dotted on the two sides of the body, with white asymmetrical rings. When these animals are upset, they have the habit to raise their wide conical tail, exhibiting the lower red or orange part, as is the case of the Cylindrophis rufus.

Squat, with a short tail, The Python curtus is not more than 2,75 m long © Dr. Giuseppe Mazza
All these serpents are excellent burrowers, less specialized, however, than the typhlopids, and are found only in soft grounds of the tropical forests, usually under pieces of trunks and vegetal debris.
They often go hunting, also outside, during the night, advancing slowly on the soil, looking for their preys. These are almost exclusively other reptilians, mainly small serpents and, in some instances, small digging saurians. For this reason, the zoological biologists call them ophiophagous and saurophagous.
It seems that they are all viviparous.
The subfamily of the Uropeltines (Uropeltinae), forms a much homogeneous and very diversified group in the same time. They are found only in the south of the Indian peninsula and in Sri Lanka, but in this area, rather small, there are not less than 7 genera and more than 40 species.
They are small or very small serpents. The biggest ones, such as the Uropeltis ocellatus and the Rhinophis oxyrhinchus, reach the 54 and the 58 cm, but most of them are under the 30 cm of length and some of them are only 10-12 cm long.
They have a conical head, not distinguished anatomically from the thickened neck, pointed muzzle, cylindrical body and short tail.
They do not have any more residual of the “pelvic girdle” and the ventral scales, even if still narrow, are already differentiated. The eye is very small, usually covered by a big cephalic plate.
The most peculiar morphological character of this group is however represented by the tail. In this organ we may in fact observe all the stages of a specialization, which cannot be seen in any other serpent. In some species, this is still of normal, conical shape and only the terminal scale is a little more ventrally flattened. But, depending on the genera, this scale may enlarge and ornate of thorns and crests, or participate to the formation of a real caudal shield, provided with a bony support.

12 m long, the Python reticulatus is nowadays the longest extant serpent © G. Mazza
The extreme stage is given to us by the genus Rhinophis, characterized by a cylindrical tail ending brusquely with an inclined oval shield.
In this case the function of the organ is clear, as it serves to close the holes burrowed by the serpent.
But the herpetological zoologists wonder if this is really the only function, and what is the meaning of the thorny scales on the tail. Ever answer, at present, may be only a guess.
In the Platyplectrurus, which are the least specialized of the uropeltines, the rostral scale is rounded; the cephalic plates are a little more numerous, the eye, even if small, is placed at a normal level, and the tail ends with a simple spine.
The behaviour and the habits of the uropeltines are little known, as is the case, anyway, of most of the burrowing reptilians.
It seems that they emerge rarely and that they are located in various biotopes, even if most of them live in the humid forests of south-west India and of Sri Lanka, and many species may reach and overcome the 2.000 m of altitude.
Their diet is based mainly, if not exclusively, on earthworms and small insects’ larvae. It seems that they are all viviparous.
The third subfamily in which is subdivided the family of the aniliids, is that of the Xenopeltines (Xenopeltinae), represented by one species only, the Xenopeltis unicolor of south-eastern Asia and Indonesia.
This unusual serpent, about a metre long, with numerous and serrate teeth which form a characteristic of primitiveness, is by sure less specialized than the other aniliids, and gets close, for many characters, to the classical colubers.
The head, flat and covered by enlarged scales, is just differentiated from the neck. The body is not regularly cylindrical on all its length, and the tail is about one tenth of its total length. The scales are broad and extremely smooth. The ventral plates are well diversified and extended, almost like the colubrids.
There is no residual of the pelvic girdle. The iridescent reflections of the skin of this animal, violaceous-brown on the back and pale on the belly, have earned it the name of “rainbow” snake.

The Liasis childreni lives on the soil, in North Australian arid zones © Dr. Giuseppe Mazza
The Xenopeltis regularly get out during the night, easily move o the soil, and if we try to seize them, show certain agility, without looking immediately to hide into the ground.
In reality, this species has semi-burrowing habits, and hunts well on the surface and also in the superficial layer of the under-wood.
It eats mainly amphibians, and, occasionally, whatever small vertebrate it may catch, serpents included.
The members of the family of the Acrochordids (Acrochordidae) have specialized for the aquatic life, and even more than the xenopeltines, they are closer, for many characters, to the colubrids. In fact, they have been classified among these last for long time and the debate about this matter is still going on between the biologists.
Without any doubt, they are the least elegant among all serpents, with a broad an flat head, not distinguished from the neck, a heavy body whose skin, too abundant, seems pleated, and a rather shirt tail. The tegument is formed, on the head too, by tiny over posed scales, all identical, and is therefore much appreciated for the fabrication of leather goods.
The adaptation to the aquatic life is evident in the nares provided with valves, located on the upper part of the muzzle, in the small eyes, also placed up, and in the absence of ventral plates. But the tail is not flattened laterally. The acrochordids are practically unable to move on the ground, and being viviparous, deliver in water.

The Morelia spilota is semi arboreal. Lives in Australia and New Guinea, reaches 2 m © Dr. Giuseppe Mazza
Only two species of them are known.
The biggest, the Acrochordus javanicus spread in the whole south-east Asia up to the north of Australia, may reach the 2,50 m of length.
This big serpent lives exclusively in the rivers, ponds and mainly in the deep marshes rich of vegetation. It bears well also the saline waters, but is totally absent from the barren coasts.
The second species, the Chersydrus granulatus, has a more extended repartition area, seen that it is found from the coast of south-western India up to the Solomon Islands, therefore under the Wallace Line and east of Papua New Guinea, that is, eco-geographically speaking, in the oceanic portion of the Australasian eco-zone.
Smaller, this serpent distinguishes from the previous one for the little less bulky body, a longitudinal keel on the belly and the presence of enlarged scales on the back.
Very little is known about its life habits, but it seems that it is found only on the seaside or in the brackish and calm waters, often not much deep. In fact, they have been observed while moving slowly in the mud, during the ebb tide, moving from a poodle to another. Preferably, in fact, they nourish of fishes.
The family of the Boidea (Boidae) is that best known to the non professional public. A relatively homogeneous group, with serpents having maintained some primitive characters, in particular some remains of the pelvic girdle. Besides a vast number of big and very big size species, found in the inter-tropical zone, they have also some small “hypogeous” (living under ground) form, especially in the arid zone of Mediterranean type.

TheAspidites melanocephalus of Nord Australia eats, rare case for boids, serpents © Dr. Giuseppe Mazza
No other group of serpents is so well represented by the fossils, which evidence, starting from the Cretaceous; species lived in Europe and America. Some of them were enormous (Gigantophis, Madtsoia), others of medium size, and in several cases are present also some typical characters of the burrowing serpents we shall find later on in the Erycines (Erycinae).
Apart these last ones, the fossils of boides disappear suddenly starting from the Pliocene, in a mysterious contrast with the previous abundance of forms.
The boides are classified in four subfamilies, well defined in base to the osteological data of the head, which however not always match with the external look: the Pythonines (Pythoninae), the Boines (Boinae), the Bolyeriins (Bolyerinae) and the Erycines (Erycinae).
The Pythonines (Pythoninae) live only in the warm zones of the Old World, from Africa to the Philippines and up to Australia.
Usually, they are big or very big serpents, with heavy body and rather short tail. The head, well distinguished from the body, shows a typical elongated muzzle, somewhat reminding that of a dog.
The greatest species are prevailingly terrestrial and live in the bushy savannahs, in the edaphic savannahs, and in the forests, quite often along the rivers, where they can swim very well. They spend the day in hollow logs or in dens built by other animals, and get out during the night for hunting. The smallest species are often more or less arboreal or branch species, and are all oviparous. The genus Python is the most common. There are three species of it in Africa, all south of Sahara. The African rock python (Python sebae) is the most spread, and may reach the length of 7 m!
It may be found in various environments, from the Sahél zone, up to the Senegal forest and the Cape, often along the banks of the rivers, but never in zones without vegetation, inside which it can camouflage more for ambushing than for defending from possible predators (at the maximum, crocodiles and lions), and animals like elephants, rhinos and buffaloes which might walk over them.

The South America Corallus caninus is almost alike Chondropython viridis of New Guinea © Dr. Giuseppe Mazza
As soon the occasion arises, it easily and willingly climbs the trees, and even if not being aggressive, when disturbed and put in conditions of inability to escape, it defends with vigour.
Nobody can keep up with the big specimens, but also the 2 or 3 m individuals may bite deeply, causing serious injuries.
The Royal python (Python regius), localized in western Africa and, especially, in the Sudanese region, is much smaller, but somewhat bulkier.
Possibly, in the wild, it does not over come the 2 m, and usually the seized specimens were measuring 1-1,50 m.
When it is disturbed, it reacts in a characteristic way, rolling the body in tight turns, with the head inside. Seen its remarkable strength, it is difficult to unwind it, and, really, it would be better tot to do this for its dangerousness in these emotional conditions, however, when in this position it may be easily transported.
Also this serpent is strictly terrestrial, even if being a good swimmer like its bigger relative.
The third species, the Python anchietae , is found in Angola and the nearby regions, but is less known by the zoological biologists.
There are no pythons in western Asia; they reappear only in India with the Python molurus.
This big serpent, which exceptionally overcomes the 6 m of length, is familiar in all south-eastern Asia and in Indonesia, and is very easy to see it in the zoological gardens and the aquatic parks.
The Reticulated python (Python reticulatus), more oriental than the previous species (it is absent in the Indies, but it extends up to the Philippines), is most probably the greatest extant serpent on the planet.

The Constrictor constrictor or Boa constrictor is the only one bearing cold © G. Mazza
It seems that some biologists have sized 9-10 specimens, and in some instances even of 12 m! Obviously, these are biological exceptions, because already meeting a 5-6 m specimen is something exceptional and in any case, impressive.
Like all pythons, it is a prevailingly terrestrial animal which nourishes of mammals of remarkable size: small antelopes, wild board and canids of various species, without anyway disdaining smaller preys such as birds and rodents.
But obviously, if annoyed or menaced, a Tiger (Panthera tigris) might easily kill this powerful animal.
The swallowing capacity of the big serpents has been exaggerated by the popular fantasies and by many adventure novels. It is zoologically impossible, that a specimen of these wonderful serpents may swallow an adult man or an ungulate of equivalent weight. After some zoologists it seems that they are able to estimate with a certain precision the volume of a hypothetical prey, and tend therefore to avoid, even if easy, the too big preys, such as, for instance, an adult man.
This is due because during the digestion they lose their moving and defensive capacities, remaining inactive. A prey too big to swallow would mean therefore more time of vulnerability towards possible predators.
On the other hand, occasionally appear in their menu animals much more fearsome to our eyes, like a 30 kg panther; but a tiger will be by sure avoided, as the African species avoid an adult lion.
Despite being so big, a reticulated python gets frequently and willingly close to dwellings and even to the outskirts of the cities, attracted probably by the abundance of rats, which live between the rubbish and perhaps, also by the presence of small dogs and barnyard animals, such as geese, hens, goats, etc. which, in the Asian countries are bred also in the cities.
Further to east, we find two other species of pythons: the Python curtus and the Python timorensis, in Indonesia only, which reach the 2,75 and the 3,50 m respectively.
The first, characterized by a particularly stocky body and the short tail, even if not being strictly semi-aquatic, seems to have an habitat well associated with the water streams. In Indonesia, and especially in Australia, there are several genera of pythons, characteristic and endemic to these zones.

The Epicrates cenchria, called rainbow boa, is one of the most multicoloured © Dr. Giuseppe Mazza
Those of the genus Morelia, similar to the classical pythons, but less massive and long no more than 4 m, live in the humid forests. They are semi-arboreal and are endowed of a prehensile tail.
The other two genera, Liasis , quite diffused from the southern Philippines and from Timor to Australia, and Aspidites, limited to Australia, differ from the other pythons due to the presence of great cephalic plates. Also, their colour is more uniform, usually brown on the back. The Liasis are terrestrial, but occupy various biotopes, both in the forests and in the savannahs or the rocky zones. The greatest one, the Liasis amethistimus, may become a giant: by sure more than 6 m, and they say that in northern Australia they have killed an 8 m long specimen! However, most of them do not overcome the 2-3 m.
Their diet is based mainly small and medium sized terrestrial mammals and some saurians. The Aspidites melanocephalus, rather slender serpent with black head and transversal bands on the body, lines in north Australia and has the particularity of being essentially “ophiophagous”, nourishing of serpents. Case quite rare in the boides, it eats indifferently poisonous or harmless species.
The Aspidites ramsayi, whose habitat stands in the arid zones of central Australia, has a more classical diet, even if with a remarkable input of saurian. Both reach, at the maximum, the length of 2,50-2,70 m, but most of the specimen is much smaller.
In New Guinea and in the York Peninsula, lives the unusual and quite fine Chondropython viridis, the only pythonine clearly arboreal.
It is a medium size serpent (from 1 to 1,80 m) which resembles in an astonishing manner to a boa, also arboreal, of South America, the Boa canina by some zoological biologists called Corallus canina.

The anaconda (Eunectes murinus) eggs open inside mother’s body © Dr. Giuseppe Mazza
Also in this serpent there is a prehensile tail and the colour of the body is pale green, with an irregular band along the spine, which is little prominent. It nourishes especially of amphibians.
There is another pythonine, the Calabaria reinhardti, which has much clearly adapted to a completely different particular type of life: very rare event in the Boidae, it has transformed in a burrower. It is a small, 60-90 cm long, serpent, with short, conical and little distinct from the neck, head, the thick tail is very short and the body is cylindrical, covered by smooth scales, some of which narrow ventral and this is quite different from what we might imagine from a python.
During the night, often, it moves around on the ground, but when disturbed it tries to hide inside the soft soil, and spends most of the day under the layer of vegetal debris. When they try to catch it, or it is disturbed in other way, it winds up quickly, forming a compact ball, like the Royal python (Python regius). This only species is found in all humid forests of western and central Africa. The diet is based on rodents, and, without any doubt, on saurian.
The subfamily of the Boines (Boinae) occupies the New World and in Madagascar roughly the same place held by the pythonines in the Old World. However, they are all “viviparous” and few of them may reach the size of the pythons.
The genus Boa (after some zoological biologists, it has been suggested during the eighties of the XX century, to replace it with Corallus, the debate about this by the International Commission of Zoological Nomenclature “ICZN” has not yet been concluded) includes several species of tropical America, of which only a small number reaches the Antilles.
They are semi-arboreal serpents, living often near water streams, without any doubt for a major abundance of preys, seen that, even if being excellent swimmers, the do not have at all aquatic habits. Contrary to what many believe, these animals are not so big. Most of them does not over come the 2 m of length, and they predate especially small rodents, arboreal opossums, birds and saurians.

The Candoia carinata, called Pacific boa, lives in Indonesia and New Guinea © Dr. Giuseppe Mazza
They have several colours, with complex drawings: for instance the Boa annulata is brown, with more or less marked in a darker tonality, the Boa cooki, is almost uniformly of sand colour, and the Boa canina is green with white dots on the back. This last species, more arboreal than the others, gets rarely down to the ground, as it nourishes mainly of birds Psittaciformes, and occasionally, of small monkeys.
We have already briefly treated about the astonishing phenomenon of “evolutionary convergence” which it presents with the pythonine Chondropython viridis of New Guinea; however, the different alimentary regimen is a sign of the fact that these two species do not occupy the same ecological niche. Even if they are little dangerous, the true boas are usually quite short-tempered animals, rather difficult to seize, and often remain aggressive also in captivity.
They have particularly long teeth (this is a morphological character we find also in other species and genera of arboreal serpents, mainly between the ornitophagi, that is those which nourish of birds) which may cause deep and painful injuries.
The Boa constrictor (Boa constrictor) is the most known of the group. Spread from northern Argentina to Mexico is usually classified in a similar genus Constrictor, that is as Constrictor constrictor. It is the only member of the family reaching the temperate zones at the northern limit of its repartition area, hibernating during the cold season.Contrary to their reputation, the biggest specimens do not overcome the 4 m of length, and most of the adults are included between the 2 and the 2,50 m.
Their behaviour is very similar to that of the true boas, but they seem to be less short-tempered and cope better with the captivity. They are often found, in fact, in the terraria-vivaria of the zoological gardens and the aquatic parks.

The Sanzinia madagascariensis is a rare boid of Madagascar © Dr. Giuseppe Mazza
The boines of the genus Epicrates, form an important group, zoologically speaking, widely diffused in the continent as well as in the Antilles, where, in several islands, are found particular species and races or subspecies
The Cuban one, the Epicrates anguilifer, reaches a size bigger than the boa constrictor, whilst the others are significantly smaller.
The Epicrates cenchria of central and southern America is often called by the zoological biologists rainbow boa, due to the iridescent reflections of the scales.
The only boa whose mass may vie with that of the big pythons, Python reticulatus, included, is the Anaconda (Eunectes murinus), living in the northern part of South America, in the Amazonian forest. It is an ovoviviparous species.
For long, till the eighties of the XX century, the zoological biologists thought that it was viviparous like the other Boinae, but later studies by means of film shoots concomitant to those of dissection done on animals (pregnant females found dead), have shown the existence of a development of cleidoic eggs inside the oviducts. During the gestation, these open at specific times, releasing already formed cubs, whereas the white fragments of the shells, with morbid consistency, are reabsorbed by the mother in order to recover part of the lost mineral resources.
From the vaginal region comes out an apt-presocial progeny, already able to creep, which effects at once the first moulting, after a few hours, and reaches in a rather short time, following precise growth curves, the allometric dimensions of the species, that is, necessary for reaching the sexual maturity.
By sure, specimens overcoming the 7 m of length have been seized, and it is possible that some big male may reach the 9 m, or perhaps, exceptionally, the 10 m! On the contrary, the numerous reports of countrymen, travellers, fishermen of those countries stating of up to 22 m long specimens, stand, to present, confined in the legends, possibly fruit of imagination, if not even of deliberate lies.

The Acrantophis dumerili lives in Madagascar and in Réunion Island © Dr. Giuseppe Mazza
A curiosity about this matter: during the second half of the fifties of the XX century, the biologists of the Natural History Museum and of the Zoological Society of New York, had made available an award of 5.000 USD for the person who should have recovered a 12-13 m specimen of Eunectes murinus, for having the same available and for being able to study its biology.
Till now, in 2011, such award has not yet been claimed by anyone ! These big serpents carry on a semi-aquatic and semi-arboreal life along the water streams of the basins of the Amazon Rover and of the Orinoco. Like many of the big saurian living in the same geographic area, they often stay on the branches which sick out on the water, also at level of the Mangroves associations, and jump in it by the first sign of danger.
It seems that the diet characterizing them differs depending on the regions. Some populations are mainly piscivorous, and others nourish mainly of mammals, birds and reptilians. In the stomach of a 7,50 m long anaconda, they have found a 2 m caiman!
In the basin of the Paraguay, there is a much smaller species, but with analogous life habits.
The only boides of the Old World are represented by two semi-arboreal genera of Madagascar: Acrantophis and Sanzinia.
Apart the unusual geographic distribution, recalling the iguanids, these animals do not have special characteristics and are very similar to the New World boas.At times, they are even classified in the same genera, Constrictor for Acrantophis and Boa for Sanzinia.
Three genera of boas of tropical America, Tropidophis, Trachyboa and Ungaliophis, form a homogeneous group and somewhat different from the othes forming the subfamily of the Boines (Boinae).

The Eryx colubrinus mainly live interred in desert zones of del North East and Central Africa © G. Mazza
They are very small serpents, mostly of the order of one metre, usually semi-arboreal.
Very little is known of many species.
The three genera are represented in the continent, and there are also many species and subspecies of Tropidophis in the Antilles.
The interesting subfamily of the Bolyerines (Bolyerinae), has two rare species in the Mascarenhas (archipelago of the Indian Ocean off Madagascar), Bolyeria multicarinata and Casarea dussumieri, nowadays exterminated in Mauritius Island, and still found, enormously rarefied, on the near Round Island.
They differ from the boines for some osteological particularities of the maxilla and of the vertebrae, and for the absence of pelvic girdle.
The are medium sized animals, from 1 to 1,50 m of length, semi-arboreal and with a look similar to that of the other small boas.
It is possible that their rarefaction depends on the disappearance of their natural biotope, mainly destroyed by the goats introduced by the man, rather than by a direct elimination.
These two species, of remarkable zoological interest for the biologists, in a few years (due to the massive decline started during the ’70 of the last century), will be probably disappeared, even before having been able to study them seriously.
The subfamily of the Erycines (Erycinae) collects a certain number of small boas, often semi-burrowers, which probably are not even relative.

The Eryx johnii lives in the arid regions of Iran, Pakistan and India © Dr. Giuseppe Mazza
Contrary to the Calabaria and to the most of the serpent living more or less hidden in the soil, they are not found in the tropical forests, but in the sub-arid zones of the boreal (northern) hemisphere.
Even if being perfectly capable to burrow in the soft ground, many are however sabulicolous (in ecology, this is said of an organism vegetal as well as animal, living on sandy soil, or immediately under it). In most of the cases, they stay in the dens of the rodents.
They get out very frequently during the night and nourish of micro-mammals and of saurian.
Like the Calabaria, they are easily identifiable for their conical head, not distinct from the neck, the cylindrical body and the short tail.
The genus Eryx is distributed from the zones of the African Sahél and the Sudan (Eryx colubrinus, Eryx muelleri) to India (Eryx conicus, Eryx johnii) and from North Africa to central Asia, including the south-eastern Europe (Eryx jaculus).
All Eryx are oviparous and have a length varying from 40 to 80 cm. On the contrary, the erycines are quite well represented in the New World.
The Charina bottae is found in the North American west, from British Colombia to California. It is not, therefore, a typical animal of the dry regions, as the Eryx, usually are, even if it has their same look and habits.
The second species of the New World, the Lichanura roseafusca is in some respects an intermediate form between the erycines and the boines. Was it not for its habitat (the arid, but not desert regions of south-western USA and north-western Mexico), we might easily classify it with the true boas.

The Lichanura trivirgata is common in south USA deserts and of Mexico © Dr. Giuseppe Mazza
This serpent, from 60 to 90 cm long, does not have the conical head and nor the extremely short tail typical of the burrowers, although these characters are slightly outlined.
Primarily nocturnal, it slowly moves on the surface of the soil, avoiding the uncovered zones and despite spending good part of its time in dens or hollow logs, does not show any tendency to start burrowing by itself. Finally, it is viviparous.
There is, then, a last group, much isolated, which sometimes is classified among the boines and, other times between the erycines, formed by small terrestrial boas of the genus Candoia (by some zoological biologists indicated as Enygrus) of north-western Oceania. These serpents, with heavy body, wide and triangular head, well differentiated from the neck, somewhat resemble to the vipers and are short-tempered.
Like some Trimeresurus (by other authors indicated as Bothrops), they lead shots with such an energy that at times they may even raise from the ground with the whole body. But at least some of them have a more peaceful defensive technique, rolling up into a compact ball, with their head more or less sunken at its centre.
Little is known about their behaviour, their habits and their alimentary ecology, but we hold some studies of the second half of the years ’70 of the XX century, about their locomotion. When they move on the sand, they perform, even if more slowly and with more uncertainty, of a gait recalling the lateral progression of the serpents of the genus Cerastes.
Infraorder of the Caenophidians (Caenophidia)

2,6 m long, the Elaphe obsoleta is the greatest serpent of Canada © Dr. Giuseppe Mazza
This infraorder includes the family of the Colubrids (Colubridae), subdivided in ten subfamilies, the family of the Elapids (Elapidae), with no subfamily, the family of Hydrophiids or sea snakes (Hydrophiidae), subdivided in 2 subfamilies and that of the Viperids (Viperidae), and subdivided in 2 subfamilies.
The family of the Colubrids includes by itself alone the two third of the extant serpents, and since always this great group is the despair of the systematic biologists.
As a matter of fact, although there is some small specialized group easy to classify in distinct subfamilies, it is extremely difficult to establish rational subdivisions between most of the species.
Unluckily, in this case, also the palaeontology was, and is, of no help, for the herpetological-zoological biologists.
The fossil colubrids are very rare in the layers of the European and North American Tertiary, and when they become somewhat abundant, in the Pliocene and in the Pleistocene, they mostly concern the present genera.
It is much possible that the group has developed and diversified in the inter-tropical zones, still little known, much before of its relevant diffusion in the temperate zones.
The classification I will follow for the family of Colubridae and, more generally, for the Caenophidia, follows a very practical criterion in order to render easier the description of this difficult and very important group.

The Elaphe longissima lives in Europe and symbolizes medicine © Dr. Giuseppe Mazza
Therefore, even if not reaching the point of separating the colubrids in several distinct families, as was done during the second half of the years’70 of last century by the British herpetological biologist of Cambridge University, Prof. G. Underwood, we shall accept, to the level of subfamilies, several of the subdivisions this author has proposed, as they are usually accepted by the International Commission of Zoological Nomenclature (ICZN).
We remind, however, that the old classification proposed by the great paleontological biologist Alfred Sherwood Romer of the Harvard University, which classified the serpents in function of the comparative anatomy of the venomous apparatus and of the presence or not of teeth specialized in the conveyance of the venom (which already subdivides them in harmless and poisonous), as well as of the position they occupy in the maxillas.
Based on this, if there are no specialized teeth and all do have the same dimensions, the serpents are defined “aglyphous” and are usually not venomous; if, on the contrary, there are teeth of conveyance of the venom, associated with the venom gland and are placed back in the maxillas, they are defined “opistoglyphous”, usually little poisonous, in any case not fatal for the man, except rare cases like the Boomslang; and if they are placed ahead, in the maxillas, they are defined “proteroglyphous”, very venomous, like the cobras, dangerous for the man; lastly, if the are hollow like the hypodermic needle of a syringe, they are called “solenoglyphous”, like the crotals and the viperids, standing among the most poisonous and fatal for the human being.
This taxonomic system, which led the American biologist to subdivide the colubrids in aglyphous and opistoglyphous, does not have nowadays any more taxonomic value, seen that the great grooved fangs have appeared independently in the various evolutionary lines. However, we have to say that along the text, I will refer to this classification only for explaining the reader if the species we are talking of is or not poisonous and, if yes, where are placed its conveying venom teeth.
From the functional and practical point of view, this scheme remains in any case very useful, even if less for the study of the Natural History of the species. Same thing may be said for the presence of a specialized labial gland, with toxic secretion, called “parotid” (nothing to do with the mammals’ parotids), called also Duvernoy’s gland, after the name of the comparative-anatomist, zoological biologist, who, in 1832 discovered it, which, in effect, is found in all the opistoglyphous, but which is however present also in a certain number of aglyphous, belonging to the most diverse groups.

The Elaphe oxycephala, known also as Gonyosoma oxycephalum, is home in Southeast Asia © G. Mazza
The subfamily of Colubrines (Colubrinae), also in the strict sense we give in this introduction, contains however about half of the Colubridae of the entire world, and in its members we may see several particular eco-evolutionary adaptations.
We shall rightly quickly review these last aspects, admitting in any case that in some instances we shall meet more phenomena of “evolutionary convergency” rather than real and proper phylo-genetic relationships.
The most classical type, that of the “colubers”, present in the school books of Biology, corresponds to the medium sized terrestrial serpents (usually 1-2 m of length), with slender but robust body, and the head often carried up (raised from the ground), well distinct from the neck. This is the case of the numerous members of the genera Coluber and Elaphe, spread practically everywhere (they are absent only in tropical Africa and in Oceania).
In northern America, the first ones are called whip snakes or racers, whilst the second ones are called rat snakes.
In Italy, the most known species of the above two genera are the “Biacco” (Coluber viridiflavus) and the Aesculapian snake (Elaphe longissima). From the mating habits of this species, was drawn the symbol of the physicians, or of Aesculapius, consisting in two serpents wound by the tail.
In the temperate regions, they are all diurnal, and actively hunt rodents, as these represent their preferred preys, to which we have to add the birds which nidify on the ground or on the low bushes, the lizards and at times other serpents, whilst the amphibians are almost never taken in consideration.

Melanistic specimen of Coluber viridiflavus © Dr. Giuseppe Mazza
In spite of being excellent swimmers, their choice habitat is represented by the thickets and the semi-covered zones.
Even if not particularly aggressive, these serpents may defend vigorously only if attacked, and by that time they bite without any hesitation. In the warm regions, some specimen of these colubers may reach remarkable dimensions.
For instance, the members of the genus Ptyas, of southern Asia, and those of the genus Drymarchon, of tropical and sub-tropical America may often be longer than 2 m.
But the record seems to belong to some Zaocys of south-eastern Asia and of Indonesia, which may reach also the 4 m.
Almost all these serpents are “aglyphous”, however there are species of this group which back of the maxilla have a great grooved fang.
This is the case of the Montpellier snake (Malpolon monspessullanus), typical of the Mediterranean region.
Numerous terrestrial colubers live in the desert zones, for instance the agile Psammophis of the Sahara, predators especially of saurian and the several Coluber, whilst in the south-east of USA we find the Masticophis flagellum.
Any coluber is capable, if necessary, to climb hedges or bushes, especially those of the genus Elaphe, which tend to do this so constantly that they might be classified as semi-arboreal.
Usually, their ventral scales have two lateral keels, more or less accentuated, which help the anchoring on the tenuous asperities of the barks.
The African colubers of the genus Philothammus, represent a good example of these little specialized semi-arboreal serpents. The Philothammus hoplogaster nourishes mainly of amphibians, and in some cases it catches them even in the water. The climbing capacity is greatly developed in the flying snakes of southern Asia, belonging to the genus Chrysopelea.

Melanistic specimen of Coluber viridiflavus © Dr. Giuseppe Mazza
Despite their agility on the branches, they do not have the same speed as many other arboreal, but they are able to climb on vertical and practically smooth trunks, of the great trees, better than any other serpent, thus occupying an ecological niche poorly considered by other ophidians.
There are also other colubrids which, even if not holding particular adaptations, apart the colours of the livery, often green or similar to a bark, live mainly between the shrubs and the bushes.
This is the case, for instance, of the American Opheodrys and also, even if in a less marked way, of some odd Psammodynastes of southern Asia, small opistoglyphous serpents resembling extraordinarily to the vipers and having two big teeth placed in the fore part of the maxilla.
The true arboreal colubers, practically localized by the zoological biologists in the more or less humid inter-tropical forests, may be easily recognized for their elongated body, in some instances almost filiform, and slightly flattened on the sides.
Most of them have keeled ventral plates. Many have also a stripe of expanded scales on the spine and lateral scales placed in slanted rows on the sides.
The green colour is extremely common in these serpents, even if they are variously sand or brown speckled, whilst others are bronze coloured with pale longitudinal lines. Almost all have a thin head, sharp muzzle and great protruding eyes.
These morphological characters are particularly remarkable in genera such as Dryophis, of south-eastern Asia, without any doubt the most specialized arboreal serpents, in the African Thelotornis kirtlandii and in the American Oxybelis.

The Boomslang (Dispholidus typus) is a nasty African opistoglyphous © Dr. Giuseppe Mazza
Nevertheless, most of the arboreal serpents have not pushed so much ahead their evolution.
Among the less evolved forms, even if already characteristic, we may remind the genera Ahaetulla, Asiatic; Dendrelaphis, Australian; Gastropyxis, African; the Leptophis, American, etc..
The genus Langaha of Madagascar has some funny nasal appendices which pose different evolution problems, about whose meaning we shall treat later on, when we shall discuss about the physiology of the organs and systems of the ophidians.
Other arboreal colubers, on the contrary, are characterized by a wide and bulky head, protruding at the extremity of a thin neck.
The Boiga, spread from tropical Africa to Indonesia and rather little specialized, have a very robust body, whilst the American Imantodes or the Malagasy Lycodryas and some other genera, are thin like the species with elongated head.
The rapidity with which these “liana snakes”, as these real arboreal colubres are often called, are able to disappear between the branches is astonishing.
They are, however, less at ease, as we have already seen, on the big vertical trunks: they are best suited, in other words, to move on a quite uneven substratum, rather than to climb vertically.
Their food is mainly based on birds and saurian; the last ones being the preferred dish for the small species. Several other species, with broad head and thin body, like those of the genus Boiga, are often ophiophagous.

Boiga nigriceps. This arboreal genus is recognized at once for its typically enlarged head © G. Mazza
Most of the arboreal colubers have large grooved teeth, placed back on the maxilla, (for this characteristic, especially in the past, they were defined as opistoglyphous), and among these ones, at least two, spread in almost all Africa south of Sahara, may be dangerous to the man.
They are the thin Thelotornis, with sharp muzzle, and especially the (Dispholidus typus): a very nice brown or saurian green serpent with robust body, which may reach almost the 2 m of length, and of semi-arboreal habits.
In both cases, the mobile maxilla may turn ahead in such a way that the great back fangs become free, thus able to perform their defensive function.
The secretion of the Duvernoy’s gland seems furthermore to be much more toxic than in other species of colubers.
Several arboreal have, this time, on the anterior part of each maxilla, one or two teeth bigger than the others, which clearly are useful for seizing the preys.
The zoological biologists have hypothesized that this type of structure should be related to the seizing of very mobile preys, like birds, but seen that same is found also in terrestrial serpents with quite different habits, it seems excessive to talk about a particular ecological adaptation.
We have to keep in mind that the arboreal colubers are much more diurnal than most of the tropical serpents.
Many of them, therefore, have narrow pupils, often horizontal, and a wide binocular field of vision.

Not only boas or pythons! This Coronella girondica chokes in its spires even rather big preys, such a lizard, and gulps them merrily © Dr. Giuseppe Mazza
The colubers give us, besides all forms of transition between the terrestrial to the specialized arboreal ones, also a progressive evolution towards animals more and more hypogeous.

The Lampropeltis getulus eats also serpents of its size, venomous rattlesnakes included © Dr. Giuseppe Mazza
A first stage is represented by serpents as European coronellas (Coronella) or the American Lampropeltis.
They are not too fast animals, with a rather heavy body, which willingly eat the serpents (ophiophagous), without disdaining the saurian (saurophagous) and the small mammals (teriophagous).
They are also provided of a remarkable power of constriction, present in the Coronella and particularly developed in the Lampropeltis.
One of the last, which in USA is often called King snake (Lampropeltis getulus), is famous for the energy with which it tears the serpents of its own size, venomous crotalids included.
The great English herpetological biologists, Prof. H.W. Parker and Prof. Angus Bellairs, respectively director and curator of the “natural history museum” of London, of the Zoology Institute belonging to the Biologic Sciences of the London University and of the “zoological garden” of London, during the sixties of the XX century, had the opportunity to record in nature a Smooth snake (Coronella austriaca) while it was attacking with great skill a European viper.
It seized it by the head, winding up around its body and then swallowed it without any difficulty, not minding at all about its poison.

Pituophis catenifer affinis (= P. melanoleucus affinis) lives in Texas, Arizona, California and Mexico © G. Mazza
In this respect, we remind that other terrestrial serpents, bigger and faster, like the South American Mussurana (Clelia clelia), reaching the 2 m of length, are also known for their ophiophagous habits.
Several colubers of this type, smaller and usually long lessthan 50 cm, such as the Southern smooth snake (Coronella girondica) of the Mediterranean regions, or the Macroprotodon and also the Duberria of northern and southern Africa, the Diadophis and the North American Storeira carry on already a very hidden life, and spend lot of their time under the vegetal debris or under the stones.
They represent a transition form with the following group.
Actually, various colubrins have become burrowers, even if not reaching the specialization level of the typhlops or the amphisbaenids (for what the saurian are concerned).
There is a remarkable variability between the forms, even more accentuated than in the arboreal. A first group includes species of medium size, provided with a modified rostral scale, still quite agile, but having the tendency to look for their food digging with more or less intensity the ground or inside the dens.
Among the least specialized, we may cite the fast Arizona and Rhinocheilus, of the south-west of USA, mainly living on the surface, the Pituophis of the temperate regions of northern America, and the South African Mole snake (Pseudaspis cana), with a heavier body.

The Rhinocheilus lecontei is found in south of USA and in Mexico © Dr. Giuseppe Mazza
These serpents nourish mainly of rodents, and spend most of their time in the dens of these last ones.
Others are sabulicolous (we remind that such are all those vegetal and animal species living on or in the sand or on soft surfaces) and predate the saurians living in the same habitats.
The Ramphiophis of tropical Africa have a pointed and protruding rostral scale. The Lytorhynchus of Sahara and the Phyllorhynchus of south-western USA, having a shield-shaped rostral scale, are more specialized and spend most of their time in the sand.
The same thing happens for another type of sabulicolous, with a thicker body, cylindrical and conical head little distinct from the neck, such as the Chilomeniscus and the Chionactis of south-western USA.
A small group, including mainly African forms, such as Prosymna and Scaphiophis, is really capable to burrow in a quite compact soil and these serpents seldom appear in surface. They are animals with a prominent muzzle, flattened and pointed, particularly hard, and with some scales (inter-nasal or pre-frontal) which have fused. It seems they nourish of invertebrates.
Finally, a last type is represented by very small serpents, almost vermiform, with smooth and cylindrical body, which live in the superficial layer of debris or under the logs, such as the Carphophis and the Cemophora of the south-eastern USA. Also their diet is based on invertebrates, often earthworms.

The Chilomeniscus cinctus spends most of time under the sand © Dr. Giuseppe Mazza
The subfamily of the Water snakes (Natricinae) has little specialized individuals which have a more or less aquatic life.
This is a group whose exact delimitation is still object of discussions, and therefore we shall consider only its most characteristic forms.
These “aglyphous” serpents are quite abundant in the temperate regions as well as in the warm and wet ones.
The genus Natrix, for instance, is spread from North America to Indonesia, but is absent in most of South America and in Oceania.
The overall appearance of the natricins is equivalent to that of the classical terrestrial colubers, with head well distinguished from the neck and the body at times little more stocky.
Their length varies from the 60 and the 150 cm. In most cases, their diet consists mainly or exclusively of amphibians and fishes, but their tendency to the semi-aquatic life is much variable.
For instance, the Ringed snake (Natrix natrix), one of the most common serpents in western Europe, lives equally well in compact grounds far away from whatever source of water, and in this case it nourishes preferably of terrestrial amphibians, and in the swamps, the reeds or on the river banks, where, in the Mediterranean region, has even fixed its preferred habitat.

The Natrix maura, which never goes far away from water, is often mistaken with a viper © Dr. Giuseppe Mazza
On the contrary, the Viperine snake (Natrix maura) and the Dice snake (Natrix tessellata), located in Europe, North Africa and western Asia, never leave the immediate proximities of the water and nourish mainly of fishes.
In North America, all the members of the genus Natrix have semi-aquatic habits, and the numerous species of the genus Thamnophis hold all the intermediate forms between terrestrial animals and those strictly connected with the water.
As for certain arboreal snakes not morphologically specialized, their type of life depends from the behaviour, and therefore it is not simply deducible from the examination of a dead specimen.
It is known, however, that in the temperate regions also the more strictly aquatic natricins in reality do spend great part of their time out of the water, both in their dens on the banks during the night, especially when the weather is bad, and outside basking in the sun.
In the entire monsoon Asia, one of the most common serpents in the rice fields is the Natrix piscator, predator, as the name says, of fishes. They are, contrary to the European and North American natricins, which rarely bite even if seized, of short-tempered and aggressive animals.

The Natrix piscator eats only fishes and is very common in Asian rice fields © Dr. Giuseppe Mazza
In the same regions, the Rhabdophis, the smallest ones, are less linked to the water and mainly nourish of amphibians.
In tropical and southern Africa, the place of the Natrix is occupied by the semi-aquatic serpents of genera Grayia, Natriciteres and Lycodonomorphus.
There are only two natricins having a greater adaptation to the aquatic life, as they have the nares placed on the back part of the muzzle: these are the South American Helicops provided with only one inter-nasal scale, and the Opisthotrophis of Asia Minor, where the prefrontal is just one.
These last ones are found especially under the stones of the mountain streams, and so they occupy an ecological niche ignored by all other serpents.
The well known subfamily of Homalopsins (Homalopsinae) forms a very homogenous group of colubers, undeniably aquatic and specialized, living in southern-eastern Asia and in Indonesia.
They are all viviparous and opisthoglyphous, with nares provided of valves placed in the upper part of the muzzle and very small eyes, turned upward. Usually, the body is much stocky, with the head little distinguished from the neck, and the tail, of normal length, is not flattened laterally.

Also Enhydris enhydris swarms in the Asian rice fields, often going out on the ground © Dr. Giuseppe Mazza
Some of these, for instance the Enhydris chinensis, and more to the south, the Enhydris enhydris, at times even abound in the rice fields, and even if not very fast, often move on the ground.
This habit is ingrained enough also in the young of Enhydris plumbea, whose diet is mainly based on amphibians.
On the contrary, the Enhydris bocourti and the Homalopsis buccata, greater, (they may reach the length of 1,15 m), and much heavier, never leave the immediate proximity of the rivers and frequent mainly the large expanses of water, often resting, however, on the soil, hiding in the vegetation or in some anfractuosities of the banks.
The genera Cerberus and Fordonia live mainly on the coasts, in particular in the wetlands and in the Mangroves.
Contrary to the other species of Homalopsins (Homalopsinae), almost all strictly piscivorous, these serpents nourish also of various crustaceans, especially the crabs, which seem to be particularly sensitive to their venom.
Along with two species of Enhydris, these genera spread up to New Guinea and northern Australia. The most specialized homalopsin is, without any doubt, the Erpeton tentaculatum of the Indochina peninsula.

Erpeton tentaculatum of Indochina has mobile appendices on the muzzle and never abandons water © Mazza
This unusual and odd serpent, long from 60 to 90 cm, is much thinner tha, the other members of the subfamily.
Its head, elongated and flat, but well distinct from the neck, is incredibly equipped with a mobile scaly appendix on each side of the muzzle.
When holding with the hand a live specimen, we remain surprised by the stiffness of its body, which has nothing to do with a serpent.
Being deprived of ventral plates, the Erpeton tentaculatum moves with difficulty on the ground and it is sure that it never gets out spontaneously from the water.
Usually, it is found in the large waterways, mainly rivers and estuaries, and at times at sea close to the coasts.
It has therefore adapted to live both in sweet and salt water.
The zoological biologists thought that it had a partially vegetarian alimentation, what should have been a unique case among the serpents, but all the specimens they have dissected had in the stomach only small fishes.
In the small subfamily of the Dasypeltins (Dasypeltinae) are grouped two colubers, which, under some aspects are very close to the colubrins, due to their much advanced ecological adaptation and for their diet based exclusively on the eggs of the birds.

The Rhabdophis subminiatus lives in monsoons Asia, eating mainly amphibians © Dr. Giuseppe Mazza
It is possible that it is only a phenomenon of “convergent evolution”, similar, in some way, to what happens about the presence of a big posterior grooved fang which, as we have seen, has appeared independently in different groups.
The most common species of this group, the Dasypeltis scaber, lives in all woody and bushy savannah zones of Africa south of Sahara.
Another species, the Dasypeltis fasciata, is, on the contrary, familiar in central African forests. The famous Indian egg-eating snake (Elachistodon westermanni) is seemingly rarer, so much that very few specimens ma be found even in the zoology museums and in the zoological gardens and the aquatic parks.
They are all rather small serpents, of 60-80 cm, which carry an arboreal or semi-arboreal life, but are not much nimble and with an appearance quite similar to the classical terrestrial colubers.
On the other hand, the Dasypeltis scaber, often looks for food on the ground.
The interesting subfamily of the Lycodontins (Lycodontinae) is represented by colubers with a much primitive structure, even if, later, they have specialized a lot. But whilst many groups are homogenous and well characterized, the group of the lycodontins still arises discussion among the taxonomic biologists, and it is difficult to be determined exactly, seen that many species represent points of transition with the colubrins and the natricins.

The flyingsnake (Chrysopelea ornata) is the colubrid best adapted to arboreal life © Dr. Giuseppe Mazza
In its whole, the family of the lycodontins, if we except the arboreal or aquatic forms, is formed by small terrestrial colubers carrying on an inconspicuous or even semi-hypogeous life.
The Lycodonts or southern Asian Wolf teeth (genere Lycodon), from which comes the name of the subfamily, represent well the same.
30 to 60 cm long, with flat head little distinct from the neck, and with thick body marked by white and black rings, they are equipped with two great full teeth, placed on the fore extremity of the maxilla, and mainly nourish of small saurians.
The Dinodon of southern China are much similar to the Lycophidion.
Also in Africa are somewhat bigger forms, such as Boaedon and Lemprophis, which vaguely resemble to the European smooth snakes.
An important group of lycodontins is represented by the numerous species of the genus Oligodon, of southern and eastern Asia.
Bulkier than the previous ones, they have an enlarged rostral scale shield-shaped, and often look for food (small saurian and several invertebrates like the myriapods: centipedes and millipedes) turning over the layer of superficial ground.
Several lycodontins with cylindrical body, the flat head little distinct from the neck and short tail, are actually semi-burrowers.

The Oligodon, with bright livery, live in south and east Asia © Dr. Giuseppe Mazza
Among these, the biggest ones, like, for instance, the Francia Francia abacura which may exceed the metre in length and lives in south-eastern USA, do live in the very soft soil of the wetlands and eat amphibians.
Due to their smooth and bright scales, with iridescent reflections, are at times called rainbow snakes, name they share with several other ophidians.
Other smaller species, vermiform at times, burrow in less humid soils.
But some North American terrestrial, like the genus Haldea, are found mainly in the tropical forests.
This is the case, among others, of the Atractus in South America, of the Miodon and of the Chilorhinophis as well as the Aparallactus in Africa, and the Trachischium in India.
In almost all, the cephalic plates are enlarged and at times merged, character we find also in several serpents living mainly in the ground, in particular in the uropeltines.
They eat invertebrates, and at times small reptilians such as Typhlops and Leptotyphlops or of small digging saurians.
The Xenocalamus, spread especially in eastern and southern Africa, where the soil is drier and harder, have an even more modified morphology, with a much elongated conical head and the muzzle formed by a huge pointed rostral scale, which greatly overcomes the narrow lower jaw.

The Boiga dendrophila is an opistoglyphous common among the mangroves of south east Asia © G. Mazza
The eyes are small, and the cephalic plates rare and very big.
These serpents, from 40 to 80 cm long, seem to eat mainly amphisbaenids.
Familiar in south-eastern Asia and Indonesia, the subfamily of Calamarine (Calamarinae) recalls some burrowing lycodontins.
They are quite small serpents, often long less than 20 cm, almost vermiform, and with the cylindrical body covered by smooth scales and short tail, cylindrical too.
The head, conical and not distinct from the neck, is covered by few and big plates; the skull is particularly rigid.
The genus Calamaria, by far the most important, is common mainly in Indonesia. The other genera, for instance Pseudorhabdion of the Indo Malay peninsula, have a similar look.
All these small burrowing serpents nourish of larvae of termites and of earthworms. The subfamily of Xenodermines (Xenoderminae) includes only four genera located in south-eastern Asia.
The most particular is represented by one single species, the Xenodermus javanicus of Malaysia and Indonesia. It is a 40-50 cm long serpent, rather slender, with the head well distinguished from the neck and long tail.
On the skull, the frontal and parietal plates are segmented and, unique character in the ophidians, the back and the sides are covered by different over posed scales, with alternation of great and small ones.

Heterodon nasicus of North America has broad and flat head, with a rostral scale for digging © G. Mazza
Though this coluber has a completely different look from that of the burrowers, it lives in the very soft and humid earth on the border of the rice fields and of the swamps, nourishing of amphibians.
The other xenodermines are bulkier: they have big frontal and parietal plates and their dorsal scales, all alike, are often slightly imbricate.
The genus Achalinus includes half a dozen of species, typical especially of southern China, living under the fallen logs, or in the surface soil, and nourish of earthworms. The other two genera, the Fimbrios of Indochina and Stoliczkaia of south-eastern part of Tibet and the Borneo Mountains, are little known. They appear to be rather similar to the Achalinus.
Three genera, well separated on the geographical point of view, are united in the subfamily of Sybinophines (Sibynophinae).
The Sibynophis live in south-eastern Asia, the Parasibynophis in Madagascar, and the Scaphiodontophis in Central America.
They are small terrestrial serpents, 30 to 80 cm long, with bulky head little distinct from the neck, provided with many teeth and a much particular mandible, being the dental articulated very loosely with the rest of the lower jaw. Little is known about their behaviour. The most common species, the Sibynophis collaris, lives in Indochina in the bamboo forests and eats saurians.
The subfamily of the Xenodontines (Xenodontinae) has three genera in the New World: the Heterodon of North America, and the Xenodon and Lystrophis of South America.

A long and agile Psammophis condanarus of Sahara © Dr. Giuseppe Mazza
These medium sized serpents, with heavy body and wide and flat head, share, besides various anatomical characters, some aggressive and showy reactions, followed by convulsions and “apparent death”.
Their muzzle has a sharp edge, the rostral scale is pointed and slightly raised, but, even if capable to quickly move the soft soil and to burrow it or to widen the dens, they are not burrowers in the strict sense of the word, and they are frequently found hunting in surface. Their nourishment if mainly formed by amphibians.
The subfamily of Dipsadines (Dipsadinae) of tropical America, and Pareines (Pareinae) of south-eastern Asia, have identical appearance and behaviour.
Though it is a possible phenomenon of convergent evolution, the debate whether to consider them as an only subfamily or two different ones is still under way.
They are all small nocturnal arboreal serpents, with a rather heavy body and flattened laterally, a very voluminous head, almost cubic, with short muzzle and big eyes.
They nourish mainly, if not exclusively, of slugs and snails, and are strictly specialized for this diet, especially for the lower maxilla, more rigid than that of the other serpents, and which ends with two big curved teeth. The pareins (genera Pareas and Aplopeltura) and the dipsadines of the genus Dipsas are the most evolved and specialized.

An Indian cobra (Naja naja kaouthia) in its defensive parade © Dr. Giuseppe Mazza
The other dipsadines are a little more alike the classical colubers, such as the species of the genus Sibynomorphus, because they do not have some typical characters of the arboreal, like the lateral flattening of the body and the enlarged dorsal scales.
The family of the Elapids (Elapidae) is not subdivided in any subfamily.
The elapids are close relatives to the colubrids, so much that some herpetologists even merge the two families; but, as it seems, it is not possible to consider them as present and more evolved descendants of the classical colubers.
Despite the almost complete absence of paleontological findings, their abundance and their diversification in Australia, show that they form an old group.
The family is localized in the warm regions, but Australia, where it spreads also in the temperate zone, and has originated in south-eastern Asia and Indonesia.
The elapids, numerous in Africa, have not however reached the Madagascar. A small group, formed by few genera but by many species, lives in tropical and subtropical America.
Some elapids are big terrestrial serpents, active and strong, resembling in the appearance and in the behaviour to several colubers like the Asian Ptyas or the Montpellier snake of the Mediterranean region.
The most famous are, no doubt, the cobras (genus Naja), due to their number and the showy defensive parades characterizing them, with the fore part of the body raising up and the spread “hood”.
The Indian cobra (Naja naja), reaching the 2,50 m, is diffused all over southern Asia, from Iran to the south of China and Indonesia.
This beautiful animal, with a colour going from the sand to the black with little paler rings, at times much faded, is much less aggressive than what normally thought, and if we consider how many people go around bare legs, without any precautions, due to the type of apparel utilized in these regions, the instances of real biting are surprisingly few.

The Hemachatus haemachatus spits venom in the eyes of the intruders © G. Mazza
Things change a lot when we try to seize them, because in some populations, the animals can spit the venom at a distance even greater than the length of their own body, targeting, as it seems, the eyes of the aggressor blinding him.
The Indian cobra likes to live in humid habitats and eats mainly amphibians, without disdaining the rodents.
The Egyptian cobra (Naja haje) familiar in the arid regions of northern and eastern Africa, as well as of south-western Asia, is not more aggressive than its oriental relative.
Two other members of the genus Naja, provided with less developed hoods, are, in the savannahs, the Black necked spitting cobra (Naja nigricollis), a very harmful spitter, and, in the forests, the Black and white cobra (Naja melanoleuca) of more pacific temper.
These three African cobras may overcome the 2 m of length and have a various diet: the Egyptian cobra prefers the mammals, the black one the amphibians.
The last two African species, Naja nivea and Naja anchietae, are somewhat smaller and are typical of South Africa.
Several cobras belonging to other genera are also present in Africa: for instance, the Hemachatus haemachatus South African, a short tempered animal which willingly spits and more efficaciously than any other member of the group, or the Desert cobra (Walterinnesia aegyptia), a bulkier but less active serpent, spread too in south-western Asia.
Finally, to end with the terrestrial elapids of this group, we have not to forget the King cobra of tropical Asia (Hamadryas hannah, by some authors called Ophiophagus hannah), the greatest extant serpent, after the boids, seen that it may reach the length of 6 metres!
Though these animals, once adult, do not practically have any foe, their bite may kill and Asian elephant (Elephas maximus).

The Egyptian cobra (Naja haje) loves the arid regions of north-east Africa and of south-west Asia © Mazza
They are always rare, phenomenon which, after some biologists, is in correlation with the typical “ophiophagous” diet they have.
Their preys are represented especially by great terrestrial colubers, like the genus Ptyas, or by young pythons, whose rather limited number clearly renders necessary a huge hunting territory, thus reducing the population density.
They say that the king cobras live in couple, and that they attack whatever intruder, man included.
These animals are not shy at all, and when the advance towards a stranger, this one usually does not hesitate in flying away.
There are a small number of elapids, similar to the cobras, which have adopted a particular lifestyle.
It’s the water cobra (genus Boulengerina) of tropical Africa: a big serpent, of sand colour with black rings, spending most of the time in the rivers, hidden in the anfractuosities of the banks, and nourishing of medium-large fishes.
It is surely a little aggressive animal, seen that the cases of biting are rare, and that the fishermen meet it often.

The bite of a king cobra (Ophiophagus hannah) may kill an elephant © Dr. Giuseppe Mazza
Other two genera, always of tropical Africa, are, on the contrary, semi-arboreal, with a slimmer body and bigger eyes than the other members of the family.
The Pseudohaje, are found only in the forest. Among the Dendroaspis (or mamba), two species live in the same habitat, whilst the third one, the Dendroaspis polylepis lives in the woody savannahs from Senegal to South Africa.
The biggest ones reach the 4 m of length, and due to their size, their speed and their aggressiveness, the mambas are by far the most feared African serpents even if they cause less incidents than some big lazy vipers.
Their venom is highly toxic, and their big fore teeth, may be straightened up with a simple rotational movement of the maxilla.
It seems that the adults are definitely less arboreal than the young individuals, and in fact, most of the times they stay on the ground, by sure due to their big size.
Actually, even if some big serpents with prehensile tail such as the boas climb the trees without any difficulty, they move there painstakingly, and it is difficult to find active and fast arboreal snakes overcoming the 2-2,50 m of length.
The serpents of the genus Bungarus (or Kraits), of southern Asia or Indonesia, ore in some way an intermediate group between the terrestrial cobras and the various coral snakes of which we shall treat later on. They are big animals, which may without any problem overcome the 2 m, with a rather heavy body, with black and yellow rings, and short tail.

The Green mamba (Dendroaspis angusticeps) is one of the most feared African snakes © Dr. Giuseppe Mazza
The spine, covered by a row of elongated scales, is oddly protruding, thus to render triangular the section of the body.
If confronted to the very nimble cobras, they seem very lazy animals.
When met they do not, in fact, try to escape or to frighten people. Sometimes, they even bury their head, round and massive, in a fold of the body.
But it’s always better not to trust, as their spring may be instantaneous, and as an old Indonesian proverb recites: “The krait never bites, but when it does so, you are lost!” The bungarus or kraits are mainly ophiophagous.
As we have already said, under the name of coral snakes are indicated many species, almost all small, with a cylindrical body rather stocky, the oval head little distinct from the neck and the short tail.
Many have rings with bright colour, most commonly red, which have originated the name.
As it can be guessed from the shape, they are animals which hide under the vegetal debris, and which have a strong tendency to burrow the superficial layer looking for saurian and small serpents as, being saurophagous and ophiophagous, these are their preferred dish.
Rather slow and little aggressive, they bite only if they try to seize them, or even if they are held too long, as may be the instance of a biologist desiring to examine their morphology.

In Indonesia they say the Bungarus fasciatus never bites, but if it does so you are lost © Dr. Giuseppe Mazza
Only three genera exist in America; they are however composed by several species.
The most abundant are the Micrurus.
A species of this genus, the Micrurus fulvius, with black, yellow and red rings, reaches the extreme south of USA (New Mexico, Texas, Arizona, Florida, and Louisiana).
The other species populate central and South America up to Argentina. Most of them are less than one metre long, whilst the greatest species, the Micrurus spixi, reaches the metre and a half.
The Micruroides, of small size, are located in the southern part of North America, starting from Arizona.
The South American Leptomicrurus have a thinner and longer body than the other coral snakes of the New World, but they hold an analogous behaviour.
The most numerous coral snakes of tropical and southern Africa, belong to the genus Elapsoidea. 40 cm to 1 m long, often with black and grey rings, the carry on a much hidden life, perhaps even hypogeous. They are often found in the termite mounds, and nourish mainly of saurians’ eggs.

An interesting horizontal bands mutation of Bungarus fasciatus © Dr. Giuseppe Mazza
The genus Elaps, from which comes the name of the family, has only two species, both South African: the Elaps dorsalis and the Elaps lacteus.
They are small, 30-50 cm long, serpents, with thin body, very nimble and active, which vigorously struggle if seized, without, anyway trying to bite.
They mainly nourish of serpents of the genera Typhlops and Leptotyphlops and of small saurian; they spend their life under the vegetal debris or the superficial layer, even if they do not burrow the ground in the strict sense of the word.
The Aspidelaps, from South Africa too, have a quite different look, more similar to that of certain Australian elapids, than to that of the classical coral snakes.
40 to 60 cm long, these animals have a rather heavy body, a short and wide head, equipped with a shield-shaped, very enlarged, rostral scale, whose protruding edges are utilized by the serpent for moving the sand or the soft earth.
Despite the fact that they are adapted for burrowing, the Aspidelaps often go around on the surface of the soil.
When alarmed, they flatten the neck, slightly raise the head, hiss rabidly and do not hesitate in biting.
The group of the coral snakes is particularly well represented in southern Asia and in Indonesia, where are located several members of the genus Maticora (by some herpetological biologists called genus Doliophis), with bright colours, often in longitudinal bands.

The small Micruroides euryxanthus is the coral snake of Arizona © Dr. Giuseppe Mazza
For instance, one of the most common species, the [Maticora bivirgata flaviceps has the head, tail and the belly of bright red colour, whilst the remainder of the body is iridescent black, with a light blue or white lateral stripe.
These serpents, whose length varies from the 50 cm and the almost 2 m, are characterized by very elongated cylindrical body, short tail and the heart displaced backwards, due to the extreme development of the venom glands, which extend up the third anterior of the body.
These are animals which live hiding, but which, as it can be understood from their size, are not at all diggers. They nourish often of other serpents, and do not exhibit any aggressiveness. As they are quite rare, little is known about their nature and about the activity of their venom, and therefore about the consequences of a possible bite. The serpents of the genus Calliophis, appear to be small Maticora (many measureless than 40 cm of length), but have normal venom glands and less bright colours, with dots, very often transversal. They are found only under logs or vegetal debris. They are active during the night, and their diet is mostly composed by very tiny serpents, especially Typhlops and Calamaria.
It is possible that some elapids typical of Papua New Guinea and the nearby islands, genera such as Micropechis, Parapistocalamus, Pseudapistocalamus and Toxicocalamus, are similar to the coral snakes, but practically nothing is known about their behaviour and about the toxicity of their venom.

In the multicolored Maticora bivirgata flaviceps the venom glands occupy one third of the body © Mazza
The fauna of the Australian elapids is so much particular that it is worthy to be treated separately.
In this continent there are no species for Viperids’ (Viperidae) family, and the Colubrids (Colubridae) occupy, in small number only, the north-eastern tropical forests.
In this way, the elapids have colonized those niches left empty and, as the marsupials have done, have diversified, thus forming that typical fauna of the Australasian and Oceanic area, under the Wallace line (after the name of the British biologist and geographer Alfred Wallace, who delineated the same).
In Australia are present not less than 30 genera, that is, more than in all the rest of the world.
Most of them are viviparous, whilst all other elapids present in the other continents are oviparous.
Strangely enough, the various types of elapids we have seen till now are little represented in Australia.
There is not an exact equivalent of the various and very poisonous African or American coral snakes, and only one species, the Pseudonaja textilis, resembles to the great terrestrial cobras.
It is a diurnal serpent, at times more than 2 m long, very active and rather aggressive, or at least bad-tempered, which willingly nourishes of other serpents.

The Aspidelaps scutatus is a South African digging elapid circulating also in surface © Dr. Giuseppe Mazza
In the north-eastern Australian forests and in Papua New Guinea, the famous Taipan (Oxyuranus scutellatus) reminds a little bit the African mambas.
Also this one is a great serpent (often up to 2 m long, at times even 3,30 m), very active, fast, provided with great teeth which the maxilla, rather mobile, allows to raise even more.
At times, even the young individuals do not show an arboreal tendency.
The taipan is much feared in the regions where it lives, and actually the consequences of its bite are usually fatal, also for the man. But there have been too many exaggerations about its aggressiveness.
Being a much mobile animal, it usually avoids the contact, and therefore is less dangerous than lazier species which hit without any notice, such as the vipers.
The great Australian venomous elapids are represented by heavier and slower species of the genus Notechis (tiger snake) and Pseudechis.
Both may flatten the neck, and often all the anterior part of the body, like cobras do, but without getting up.
The Notechis scutatus, even if not semi-aquatic, is often found in the humid locations, something like our Ringed snake (Natrix natrix).
On the contrary, the Notechis ater loves the dry locations. These animals are diurnal, have a mixed diet and do not overcome the month and a half of length.

The famous Australian Taipan (Oxyuranus scutellatus) has same bad character as the mamba © G. Mazza
But there is a particular insular race or subspecies which may reach the 2,40 m. And this is not its only particularity, as the adults live almost exclusively at the expense of sea birds: the young of the shearwaters and petrels, which build their nests on the ground.
By the hatching of the eggs, they have to eat to fill up their belly, but this occurs only during a short time of the year.
Then, they find only small saurians, just sufficient for furnishing the newborn serpents the indispensable nourishment.
The two most common species of Pseudechis (Pseudechis porphyriacus and Pseudechis australis) are also of big size, as they reach the 2 m of length.
These serpents, mainly nocturnal, have venom less toxic than the members of the genus Notechis, and are often ophiophagous (particularity common to most of the Australian elapids, but the taipan and almost all the Notechis).
Many species, obviously, replace the missing colubers: that is, they have their look and behaviour, with a reduced toxicity, by sure still efficacious on the preys, but which seems to have a more defensive form.
The different species of the genus Demansia, whose size varies from 1 to 2 m, resemble much to the great and lively terrestrial colubers of the genera Elaphe and Coluber.

The Austrelaps superbus is a much venomous serpent of Australia and Tasmania © Dr. Giuseppe Mazza
Like these ones, they are diurnal, and defend themselves with energy.
The type of the less active terrestrial colubers, of medium or rather small size, like our European smooth snake, is well represented by various species of the genus Denisonia, or by the members of the genera Brachysoma, Glyphodon, Drepanodontis, etc., which nourish mainly of small saurians and are almost completely harmless for the man.
Some Australian elapids, belonging to genera Cryptophis, Lunelaps, and Parasuta, of look similar to the previous ones, with a length of 40-90 cm, have an even less showy life, as they hide, during the day, under the vegetal debris or stones. Also for them, the preferred preys are small saurians.
Some very tiny species, 15-40 cm long, has become semi-burrower, with the head not distinguished from the neck, the cylindrical body and the short tail.
The genera Melwardia and Rhinelpas are, still now, little specialized, with a round head, very little flattened; the Narophis hold already an elongated and reinforced muzzle.
The Brachyurophis are typical sabulicolous burrowers, very characteristic, with the rather heavy body and the conical head, like the Chilomeniscus of the south-western USA.
The smallest among these serpents eat mainly invertebrates, whilst the others nourish also of saurians. A last species, the Vermicella annulata, represents a rather particular type of burrowers, with a small head and very elongated cylindrical body. It seems it nourishes only of Typhlops.

Even if not aquatic the Notechis scutatus is always found in wet locations © Dr. Giuseppe Mazza
When disturbed, this serpent takes a strange defensive position, with the head leaning on the ground.
It does not raise the tail, like most of the small serpents and the semi-burrowers do, but a wide ring of the body.
Among the Australian elapids, we find very few arboreal forms, perhaps because the forest stretch mainly in the north-eastern part of the continent, in a zone where colubrids live and in particular many species which have this style of life, like those belonging to the genera Dendrelaphis and Boiga.
It is furthermore quite odd to see that the only one semi-arboreal genus of the elapids, the Hoplocephalus, occupies the outskirts of this zone, to the south and in the mountains to the west, in still luxuriant, but already cooler, forests.
These are 75 to 150 cm long serpents, with bulky head and little narrowed body, compressed laterally and with keeled ventral plates.
Being relatively little specialized, they frequent also the rocky zones, the stony grounds, and have an eclectic alimentation.
Like many arboreal, they are easily irritable, and even if not really dangerous, their bit may cause serious damages to man. Two genera of medium size (from 50 to 75 cm) terrestrial elapids, Aspidomorphus and Brachyaspis, have a rather heavy body and the head widening backward, clearly distinct from the neck.
They are harmless and greedy eaters of frogs.

The Acanthophis antarcticus seems a viper and its bites are fatal in the 50% of cases © Dr. Giuseppe Mazza
The Acanthophis, are different, just larger but with fearful bite. An example is the Acanthophis antarcticus, with bronze-brown livery and polygonal black maculae distributed in a rarefied way.
These animals correspond exactly to the terrestrial Viperids (Viperidae) and have also their appearance, with a wide triangular head, well distinct from the short but very thin neck, a much heavy body, dorsoventrally flattened, and a short and bulky tail, which is not any more utilized for the locomotion.
Very lazy, but ready to bite without even trying to flee away, they haunt, in ambush, saurian, birds and small mammals, at times partially hiding under the sand. When excited, their tail moves a lot and straightens up.
As the proximity of a prey is sufficient for trigging such a behavioural status, it is not excluded that this is one of the very rare cases of a real use of bait.
In fact, some Australian herpetological biologists have proved that birds and saurians react, intrigued, to this movement, getting so much close to be seized.
Among Australian elapids, the Acanthophis are probably the most feared both for the toxicity of their venom and because it is difficult to see them and so, they hit without notice. On the other hand, the name “death adders” is clearly allusive.
Family of the Hydrophiids (Hydrophiidae), sea snakes, has two subfamilies: that of the Laticaudins (Laticaudinae) and that of the Hydrophiins (Hydrophiinae). The laticaudins, equipped with two still wide ventral plates, are capable to move on the ground, and at least one genus, being oviparous, is obviously obliged to do so.

The Laticauda colubrina reproduces on the ground and has a very vast distribution © Dr. Giuseppe Mazza
The Laticauda, main genus of the Laticaudinae, are rather big serpents, 80-180 cm long, still not much specialized. The head, small and round, slightly distinct from the neck, has lateral nares. The elongated body is semi-cylindrical, provided with ventral plates as big as half of their length. Only the tail, quite short, tall and very flattened on the sides, reveals their lifestyle. The typical colouration is characterized by black and yellow or blue rings, regularly alternated.
Of the four extant species, two (Laticauda laticaudata and Laticauda colubrina) have a very ample distribution, from the Bengali coasts up to the Tonga Islands and of southern Japan, and then we find them also in Tasmania and in New Zealand. The greatest, the Laticauda semifasciata, lives only in the Molucco Islands, in the Philippines and in the Ryukyu; the smallest one, the Laticauda schistorhynchus, is located in the Fiji and Samoa Islands. In New Caledonia during the seventies of the last century, the zoological biologists had the opportunity to study the Laticauda laticaudata and mainly the Laticauda colubrina, for the first time in their natural habitat without interferences, during the austral winter. In this season, these serpents meet in big number: several hundreds and at times thousands of individuals on small sandy islets, within the lagoons.
It looks like that each serpent spends several weeks on the ground, where it may digest and moult, and then stays in the water for a very long hunting period, which takes it even many kilometres far away, at times up to the “Great Barrier Reef”! Mating takes place in spring, after which the Laticauda disperse and spend much time at sea, coming back, from time to time, on the sandy aits. It is here that, during the summer, the females lay their eggs in holes dug in the humid sand, above the high tide line.

The Laticauda schistorhynchus is common in Fiji Islands and Samoa © Dr. Giuseppe Mazza
When on the terra firma, the Laticauda move very little. They do not hide at all, limiting, during the warm hours, to stay in the shade, under blocks of dead coral or in the fissures of the small crest of sand which goes along the line of the high tide. Strangely, it seems that these animals do not know the danger: in fact, they never try to escape, do not flounder and do not bite when seized.
On the contrary, when in the sea water, their behaviour is quite different: they are much more active, they plunge as soon as they sight a boat approaching, and, after the local fishermen, they are never caught without reacting. They nourish only of fishes, especially eels and morays which they seek exploring the anfractuosities of the rocks and the corals.
The Laticauda semifasciata, the greatest species of the genus, particularly abundant in the Philippines, also forms crowded colonies, this time in calcareous grottoes and is, unluckily, object of systematic exploitation by man, for its skin and flesh. One fishing station has reported 10.000 serpents per year just itself and this may give an idea of the massacre. Seen their exigencies, the Laticauda are in fact easily found.
The seven species of the genus Aipysurus, with lengths of 0,80-1,70 m are already more specialized. The heavier body, the longer and flat tail, the nares placed on the upper part of the muzzle, and the ventral plates wide only one third of the body, show a more marked and stable evolution towards the sea life. The Aipysurus eydouxi is spread from the Gulf of Siam to Papua New Guinea and the north-east of Australia, whilst the other six species are found only in the eastern half of this zone, which some occupy not even its entirely.
We have little information about the biology of these animals, which rarely land and seem to be viviparous. The same remarks apply for the similar genus Emydocephalus, with two species, located one on the coasts of Taiwan and Ryukyu, the other in Melanesia.

The nares of the Hydrophiinae close tightly and the vision is perfect © Dr. Giuseppe Mazza
The subfamily of the Hydrophiins (Hydrophiinae) gathers sea snakes more specialized than the Laticaudins (Laticaudinae), but at the same time more primitive for several osteological characters, which lead to think that they have parted before the elapids. Also here the body is often more or less flat laterally, the tail is quite long and the ventral plates are reduced or not differentiated.
They are all viviparous animals, almost completely unable to move on the ground, even for short distances.
The newborn are delivered in the water, and being a precocious offspring, they are already able to swim. It is not unusual to find these serpents, masters of aquatic camouflage, tangled to the kelp (brown algae with laminar thallus) and to other seaweeds: a manner for camouflaging from the predators and surprising the preys. While moulting, as soon as the skin is lost, they are vulnerable to the sight of the predators, and same thing applies to the newborn.
But one exception only, these serpents are found especially near the coasts, isolated at times, other times in very numerous groups. They nourish only of fishes, but apart this, very little is known about their behaviour, and, mainly, we ignore the meaning of their individual displacements. South of the Equator (austral hemisphere), the delivery happens usually in November-December, in the northern hemisphere (boreal hemisphere), in February-April, but there are remarkable variations, not so much from one species to another, but among the different populations.
The Hydrophis semperi, clearly much similar to the Hidrophis cyanocinctus, populates a volcanic lake in the Island of Luzon (northern Philippines), connected to the sea by means of a narrow passage, about ten km long, with a current at times rapid. It is evident that the fresh water doesn’t disturb the sea snakes, which often venture in the estuaries and in the great rivers. Some of them have been even seized in the Tonle Sap, the great lake inside Cambodia, where flows, during the dry season, a part of the water of the Mekong River. Another species, the Pelamis platurus, is decidedly pelagic, and is found from Madagascar to the western coasts of Central America. Among the hydrophiins, in spite of their common characters, animals of different type have evolved.
Some genus, for instance Kerilia or Praescutata, has still rather classical proportions, but many of the sea snakes are massive with head little distinct from the neck and a smooth body, slightly arched towards the ventral side. The Lapemys and especially the Enhydrina schistosa, with heavy body surrounded by a wrinkly skin with a very weary look, remind somewhat the acrochordids. Another type is represented by the several species of Hydrophis and by the Microcephalophis.

The back drawing does so that this Hydrophis elegans is little visible from above © Dr. Giuseppe Mazza
These serpents have a very small head, well distinguished from the long and thin neck. More backward, the diameter of the body increases progressively, and the back side, with the tail, is huge compared to the rest. The mechanical advantages of this structure are clear: thanks to the small head and the remarkable inertia of the back, these serpents may easily beat the water, as it should be air, thing not happening with the other hydrophiins.
The Pelamis platurus is finally endowed of a great elongated head and of a much laterally flattened body, starting from the neck to the tip of the tail, with the curvature of the ventral face particularly accentuated. Most of the hydrophiins have a size included between the 80 and the 130 cm of length.
The biggest ones, the Hydrophis cyanocinctus and the Hydrophis spiralis, reach the length of 2,50-2,75 m, but there are species, such as the Astrotia stokesii, which even if not overcoming 1,60 m, are much heavier. The 60-80 cm Pelamis stand among the smallest sea snakes
All the hydrophiins have the ventral face of the body much paler than the dorsal one, and many are more or less ringed. The liveliest coloured is, without any doubt, the Pelamis platurus, black on the back and the head, bright yellow on the flat sides and on the belly. The presence of a ventral surface paler than the dorsal one, has an eco-ethological meaning, and is a phenomenon defined of “pigmentary polarity”, present almost everywhere in the animal kingdom, in the group of fishes, as well as in the class of the birds, the amphibians and the mammals. Its most salient expression stands however in the marine organisms (fishes, reptilians, mammals), as the presence of a dark dorsal surface melts (especially for the pelagic life species) with the surface of the water, which, offshore, assumes a blue-dark green colouration.
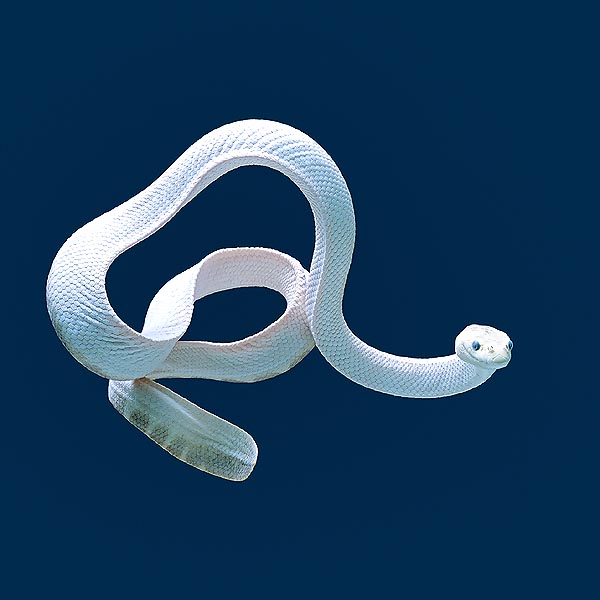
But seen from below the animal is pale and blends with the light penetrating the water © Dr. Giuseppe Mazza
This renders impossible to see the animal from the surface, whilst the pale ventral surface, which mixes with the light penetrating in the water, does so that the animal, when seen from below is assimilated to the environment, thus camouflaging from preys and predators.
Most of the hydrophiins live I the region extending from Malaysia to the Solomon Islands and from Celebes to northern Australia. Many species are located only in these places, and it seems that some of them are even limited to a small group of islands or a coastal zone.
There are however many sea snakes having an ampler distribution, east and, more often west. They are still numerous on the coasts of the South China Sea and especially of the Gulf of Siam, much less, on the contrary, in the Bay of Bengal. There are only 10 species passing to the west the level of Sri Lanka, but seven of these reach the Persian Gulf.
They are the Praescutata viperina, the Enhydrina schistosa, the Hydrophis spiralis, the Hydrophis cyanocinctus, the Hidrophis ornatus, the Lapemys curtus and the Microcephalophis gracilis.
Mainly pelagic, the Pelamis platurus has a zoogeographic distribution very vast, which includes all Indian Ocean and most of the Pacific, up to more than 40° North (Sea of Japan), or South (Tasmania).
However, it is found in the western coasts of Pacific only at the level of Central America, and it seems that it has never entered the Atlantic Ocean (too cold) in spite of the existence of Panama Canal. Finally, they are very venomous serpents; their bite may cause by sure the death of an adult human being, especially when, as it seems, they attack in group.

Melanistic reddish with drawing specimen of Vipera berus © Dr. Giuseppe Mazza
The family of the Viperids (Viperidae) is subdivided in two subfamilies: that of the Viperines (Viperinae) and that of the Crotalines (Crotalinae).
We have to say at once that to these belong some of the most venomous and mortal species of serpents of the suborder of the Ophidians (Ophidia), also for the human being.
Even though the venom apparatus of the viperids is by sure the most sophisticated among the serpents, the family has also some rather primitive characters, especially for what concerns the vertebrae and the hemipenes.
It is sure, phylogenetically, that these serpents come from quite primitive colubrids, or from pre-colubrids and not, as often thought, from evolved elapids.
Unfortunately, the paleontological findings do not furnish precise information about this subject, as the oldest known fossils (European Oligocene), belong to animals which already were very characteristic viperids.
In the whole, the present forms are of medium or small size, with the head widening backward, well distinct from the neck, whilst the body is rather heavy and the tail is short. Most of them are viviparous (the word viper, is the contracted form of viviparous) and terrestrial, in some case arboreal, rarely semi-aquatic.
The Viperines (Viperinae) are spread in most of the Old World (they are absent only in Madagascar and Oceania) and in the boreal hemisphere they occupy the whole temperate zone, with a species almost going as far as the level of the Arctic Circle! They are numerous in Africa, where probably is located the dispersion centre of the subfamily. There are still a quite remarkable number of them in Europe and western Asia, whilst only one species reaches the Asian south-east and Indonesia.

Small, with several subspecies, the, la Vipera ursinii presents primitive characters © Dr. Giuseppe Mazza
The genus Vipera has the particularity of having differentiated in the temperate zone, more precisely in southern Europe. It is possible that the various Ice Ages of the Quaternary have covered an important role in the forming of the present species, isolating, on various occasions, more or less numerous populations.
It follows that the present systematics of the group is quite complex.
However, by doing some examples, it is possible to distinguish sufficiently homogenous groups, at different stages of the evolution process. The most primitive group (corresponding to the old genus Pelias) is characterized by the little divided cephalic plates, and by the head scarcely widened backward.
It includes the Common European adder (Vipera berus), spread in the temperate forests, from the Atlantic to the Pacific and in Scandinavia almost up to the Arctic Circle, and several similar forms localized in the mountains or the steppes delimiting to the south its repartition area.
Some of these, rather small (from 35 to 45 cm of length), seem particularly primitive, and are often united under the name of Vipera ursinii (Vipera ursinii ursinii, in France and in some Italian location of the Apennines and of the Southern Alps, Vipera ursinii macropos in the Balkans, and Vipera ursinii renardi in the Romanian steppes to Altai).
Others generally considered as simple races or subspecies of the Vipera berus (Vipera berus seoanei in the north-west of Spain, Vipera berus bosniensis in the Balkans, Vipera berus kaznakovi in the Caucasus), form, on the contrary, the transition forms towards the following group.

A Vipera aspis camouflaged in the wood marginal vegetation © Dr. Giuseppe Mazza
This includes three greater species, between the 70 and 90 cm of length, characterized by a more marked division of the cephalic plates, the raised muzzle, and the head widened backward.
These characters, already clear in the Vipera aspis of France and Italy, accentuate in the Vipera latasti of Spain, Portugal and North Africa, and even more, in the Vipera ammodytes of the Balkans and west Asia.
Finally, the third group, quite separated from the previous ones, is represented by great and heavy vipers, more than 1 m long, with very wide muzzle and head, fully covered by small scales.
The four species presently recognized do live, all of them, in Mediterranean type zones, more or less arid, placed south of the habitat of the other members of the genus.
They are the Vipera mauritanica of North Africa, the Vipera lebetina, common from the Cyclades to western Pakistan and Uzbekistan, the Vipera xanthina, present in Asia Minor, and the Vipera palaestinae of the Middle East.
The relationship between the great Russell’s viper (Vipera russelli) and the previous species are not clear at all.
Although its morphology, despite the diversity of the muzzle, reminds the Vipera lebetina, its geographic distribution, primarily tropical (India, Asian south-east and Indonesia) is aberrant. On the other hand, it is the only viperid of these regions.

A Vipera ammodytes with cub, easily reconizable for the horn on the muzzle © Dr. Giuseppe Mazza
It is a nocturnal animal, located in the sparse forests or in the shrubby savannah, of which many populations are known, often quite isolated between them.
Other quite particular viper, very different from the previous one, the Azemiops feae, is located in the strip of south-eastern Tibet.
The few specimens known are coming from the mountains of Burma, the Tonkin and the south-west of China.
Considering the big cephalic plates, the head little distinguished from the neck and the short fangs, this 60 to 80 cm long serpent, with thin black rings, is even more primitive than the Vipera ursinii and occupies an isolated position in the viperids.
The warm desert zones extending from the Atlantic to the western extremity of India, are occupied by several tiny vipers belonging to different genera, but having the same behaviour and a quite similar appearance: the head is wide, well distinct from the neck, the body is bulky with strongly keeled scales and quite often placed in oblique rows on the sides.
They are all nocturnal, and spend their days sunken in the sand, leaving out the head only. Furthermore, they move with a particular locomotion technique, with lateral undulations, leaving on the sand, unmistakable traces of their passage.

The Vipera xanthina, of Asia Minor, usually warns, hissing, before biting © Dr. Giuseppe Mazza
The most common is the famous Saharan horned viper, called also Horned desert viper (Cerastes cerastes ), so called due to its long sharp scales placed over the eyes.
It’s an animal living mainly in the rocky zones, even if it may be met also in different type biotopes, from the Algerian highlands up to the Sahél zone in tropical Africa. It nourishes of saurian and rodents. A similar species, little smaller and hornless, the Cerastes vipera, lives also in the Sahara, where is found only in the great sandy dunes (structures called “ergs”, even up to 200 m tall).
Here, its colour melts, by means of a process called of “somatolysis” (that phenomenon for which the presence of a coloured livery with particular decorations and drawings, coincident with the shades, the lights and the colours of the environment forming the background, determines a chromatic condition, “without solution of continuity”, camouflaging the animal in the habitat. Just think to the mantle of the tiger or the livery of a zebra), with the colour of the substratum, grey or salmon pink as the cases. It eats almost exclusively saurian: especially the so-called sand fishes and the Acanthodactylus.
Contrary to most of the terrestrial serpents, it does not seem that these two species are territorial. During the night they may cover several kilometres, but not during the cold season, when the displacements are much limited.

Equally dangerous, Vipera lebetina is spread from Cyclades to western Pakistan and Uzbekistan © Mazza
In the erg (the dune sandy structure), cooler, the Cerastes vipera spends a period of real and proper lethargy, whilst the Cerastes cerastes becomes diurnal and sedentary on the rocks, basking in the sun during the day and hiding during the night in some deep anfractuosity.
These serpents are quite aggressive and are cause of several mortal incidents, as because they enter the camps during the night and also because it is easy to walk over them when they are interred into the sand.
We know little about the habitts of the other desert species, genera such Pseudocerastes and Eristocophis living in the south-west Asia, from the Mediterranean to western Pakistan, but it is possible that they are much alike the cerastes. Oddly enough, the Eristocophis macmahonii, which looks for the rest particularly well adapted to the sabulicolous life, holds laterally keeled ventral plates, as it the case of the arboreal serpents.
On the contrary, the Pseudocerastes do not have oblique rows of scales along the sides and get a little closer to the genus Vipera.
A last species, the Echis carinatus has several characters in common with the previous serpents, but it lives in a less arid region.

The terrible Vipera russelli, kills many in India e in south eastern Asia up to Indonesia © Dr. Giuseppe Mazza
Its quite vast range includes a narrow stripe of territory at the northern limit of Sahara, the Sahél zone and good part of the Sudanese region, besides the corresponding regions of south-west Asia and the arid parts of India up to Sri Lanka.
This small viper, very aggressive and with a strongly toxic venom, is really dangerous and deadly for the man.
All other viperines live in tropical and southern Africa, where they have evolved in different forms.
The Causus are small oviparous terrestrial vipers, rather primitive, with great cephalic plates and little heavy body.
Of nocturnal habits, they are often endowed of venom glands extending inside the neck, and mainly eat terrestrial amphibians.
The genus Bitis is one of the most significant, for its much advanced evolution and for the strong diversification it has met during the elapsing time.
Several species, rather big, represent the perfected type of the heavy and bulky viperid, with flat head, much enlarged backward and well distinct from the neck, an extraordinarily thick, and a short and thin tail which even does not adhere any more to the soil. The venom glands are enormous and the conveying teeth are longer than in any other serpent.

The Cerastes cerastes is very dangerous as it hides in the sands © Dr. Giuseppe Mazza
The most diffused species, the Puff adder ([Bitis arietans), lives in the savannahs from Senegal to North Africa, and there is also a small residual population north of Sahara, in south-western Morocco.
The other species do live in the jungle and are remarkably coloured with complicated drawings which include an astonishing variety of colours, from black to yellow and from red to blue, passing through various tones of violet and green.
The whole, in spite of its mass, renders the animal really little visible on the soil of the under wood.
It’s a classical example of “somatolysis”.
The greatest species is the Gaboon viper (Bitis gabonica ), which reaches the 1,80 m of length, and which is one of the most dangerous vipers in the world, with an extremely toxic venom.
This last is at the same time, vasotoxic, haemotoxic, and cytotoxic, properties which cause internal haemorrhages, with copious spills of blood from the oral and anal cavities, from the ears and the eyes, heamolysis, with destruction of the red blood cells and drastic drops in blood pressure, and necrosis and gangrene of the tissues in the points where the bite occurred.

The Echis carinatus is very aggressive, with a particularly toxic poison © Dr. Giuseppe Mazza
And we must not forget that the venom conveying teeth of this solenoglyphous species are so big, that the bite itself produces a very strong pain, which may be compared to that of a leopard.
It is a much reserved animal. When disturbed, it begins to emit puffs and low noises much pronounced and persuasive.
If the annoyance persists, it attacks like a lightning, with its powerful bite.
The Bitis nasicornis a little smaller, is equipped with two horny appendices which rise vertically at the extremity of the muzzle.
These solid vipers nourish mainly of rodents and are rather lazy, at least till the moment they decide to hit.
Despite being little aggressive animals, to such a point that they can be manipulated without reaction, they cause several serious and fatal incidents.
As a matter of fact, they are not at all inclined to escape or to manifest a warning behaviour, if not with puffs, when they reach the limit of tolerance, as said before.
In South Africa, several tiny Bitis clearly occupy the place of the cerastes and similar forms of North Africa and western Asia.
The Bitis peringueyi, 25 cm long, lives in the sandy dunes of southern Namib, and moves quickly with lateral undulations.

The Causus resimus is an oviparous, primitive viper. The venom is little active but abundant © G. Mazza
The Bitis paucisquamata and the Bitis cornuta, which are somewhat bigger and less specialized, occupy different zones of the African south-west, preferably sandy, on the contrary, the Bitis atropos seems to be localized in the rocky and mountainous regions of the African south-east.
All these smaller Bitis nourish mainly of saurians.
The Bitis caudalis, 40-45 cm long, frequents different biotopes, both of sand and of rock, and is by sure the most common viper in all southern Africa.
The Atheris, of the forests and the shrubby savannahs of tropical Africa, are arboreal vipers much similar to the Bitis, but with a more uniform look.
Their body is less stocky, the head is very wide and well distinct from the body, the scales strongly keeled and the tail is prehensile.
Like all the arboreal viperids, their way of moving among the branches reminds in some way that of the boides.
They are, that is, animals which move rather slowly, keep the hold using their prehensile tail and stay motionless in ambush for long hours.

The elegant drawing of Bitis gabonica is extremely camouflaging in the vegetation © Dr. Giuseppe Mazza
They are quite small (from 50 to 80 cm of length) serpents, green or variously spotted of yellow on dark background, nourishing of various preys, mainly amphibians and have an irritable character.
Their bite may have only exceptionally lethal consequences for the man.
Two rare small species, terrestrial or semi-arboreal, belonging to the genera Hindii, of Kenya and Supercilliaris, in Tanganyika, once considered as belonging to the genus Vipera, are now classified in the Bitis, even if, after some zoological biologists they are rather anomalous species of the genus Atheris, like, for instance, the Atheris squamigera .
A third African viperine, the Adenorhinos barbouri, diverges even more from the common type, for its diet, formed in good part by invertebrates and mainly earthworms, and also foe a minor segmentation of the cephalic plates, probably in relation to its diet and connected with its habits of semi-burrower serpent.
Then stands all the group of African vipers, at times united in a particular subfamily carrying on a really underground life.

The Bitis nasicornis is not aggressive, but camouflages, doesn’t escape and so is much dangerous © Mazza
These serpents, united in the genus Atractaspis, are already rather specialized burrowers, found not only in the soft earth of the forest, but also in the more compact soils of the savannahs. Strangely, one species, the Atractaspis engaddensis lives in Sinai and in southern Israel.
All these vipers, long from 40 to 70 cm, have cylindrical body and the tail rather short and thick, typical of the burrowing serpents. The head, almost not distinguished from the neck, is characterized by the rather long muzzle, with an often sharp rostral scale and the small eyes, and is covered by big scales. The Atractaspis nourish mainly of small underground reptiles, such as Typhlops, Leptotyphlops, of saurian, such as the amphisbaenides, and many Feylinia, basically, they are ophiophagous and saurophagous. Contrary to what we might expect from their type of life, these animals have teeth proportionally longer than those of many other viperids. Clearly, these are not utilized under ground, and in surface the Atractaspis hit most of the times without opening the mouth, with a lateral and anteroposterior movement of the head, digging one tooth only. The venom seems to be little toxic.

The Crotalus horridus is the terror of Canadian woods and of eastern USA © Dr. Giuseppe Mazza
The subfamily of the Crotalines (Crotalinae) differs from that of the Viperines (Viperinae) only for the presence of aparticular sensorial organ, placed between the eye and the naris, which has the same function connected to the thermal sensitivity as the labial dimples of the Boids (Boidae), and called “thermal dimples”.
They enable to “see through the heat”, to perceive the presence of a prey or a predator thanks to the heat emitted by the body, carried by the blood flowing inside the same.
The geographical distribution of the two subfamilies of the Viperinae and of the Crotalinae is complementary. The crotalines are located mainly in the New World, where by sure their differentiation and specialization have happened evolutionally speaking, and in eastern and central Asia, all regions which are very poor of viperines.
The crotalines, include two well distinct groups of animals: the rattlesnakes (genera Crotalus and Sistrurus) and the rattleless serpents (genera Trimeresurus, Lachesis and Agkistrodon).
Each of these groups, divides itself in species equipped with great cephalic plates, a character which is considered more primitive (genera Sistrurus and Agkistrodon) and in forms whose head is wholly covered by small scales.

Crotalus viridis lives the great south western plains from Canada to eastern USA and Mexico © G. Mazza
Like the viperines, the crotalines are essentially terrestrial serpents, with a certain number of arboreal species.
None of them, is however a burrower (they do not hold such primitive structures), whilst there is a semi-aquatic form.
But very few exceptions, (Lachesis mutus or muta, and also some Agkistrodon and Trimeresurus, terrestrial of South-eastern Asia), the crotalines are almost all viviparous.
But one species, the rattlesnakes are located only in North America, especially in the southern part of the USA and in Mexico.
The genus Crotalus is by far the most important.
The size of the various species has a remarkable variability (from 40 to cm to 2,40 m), whilst the overall look is quite uniform. It’s always matter of serpents with a heavy body, triangular head well distinct from the neck, and the tail ending in a rattle of variable size.
All Crotalus are mainly mammals predators and strictly terrestrial.
But the different species occupy various ecological niches, depending on the climate, the vegetal mass and the nature of the substratum.
The northern species, the Timber rattlesnake (Crotalus horridus ), well known to the early English settlers, lives in the deciduous forests of all north-western USA, and from the Canadian boundary up to the Gulf of Mexico.

The Crotalus molossus is common in southern USA and in Mexico © Dr. Giuseppe Mazza
The Prairie rattlesnake (Crotalus viridis) is, on the contrary, typical of less covered regions, in particular of the great plains of the centre of the continent, up to southern Saskatchewan and the highlands located between the Rocky Mountains and the Pacific.
Like the horned vipers, it seems that during the summer it is more or less erratic, even if keeping faithful to the deep shelters where it spends the winter hibernating.
In this immense geographic region, several races or subspecies have differentiated, so this rattlesnake is one of the serpents having a very great variability of appearance.
In the thick tropical or subtropical forests, live few rattlesnakes, among which the Crotalus adamanteus of Florida, reaching even the 2,10 m of length, and the Crotalus durissus little smaller.
The last one, spread all over Central America and most of South America east of the Andes, is rightly feared due to the toxicity of its fatal venom.
Both species are not exclusively of forest, and are found also in the glades and in the shrubby zones (bush), but their habitat is limited to the humid and warm zones.
However, apart these exceptions, the rattlesnakes are always guests of decidedly Mediterranean climate zones, more or less arid, which occupy the whole south-west of USA, from California to Texas, and most of Mexico.

The Crotalus adamanteus of Florida may be 2,10 m long © Dr. Giuseppe Mazza
Out of 27 known species, 23 are located in this type of region.
The famous Western diamondback rattlesnake (Crotalus atrox) may overcome the 2,40 m of length, and is therefore the greatest member of the genus Crotalus, followed closely by its congener, th Crotalus adamanteus, autochthonous of Florida. But it’s a somewhat rare animal, which causes few incidents.
The smaller species, such as the Crotalus pricei and the Crotalus transversus, both Mexican, do not overcome, respectively, the 48 and the 58 cm of length.
As a general rule, in the alimentation of these species, the saurian have a more important role than what happens for the bigger rattlesnakes, which, as already briefly told, are preferably theriophagous.
Whatever their size, these reptiles love staying in the bushy and uneven areas, spending their days in the dens of the rodents or in much hidden shelters, from which they get out preferably at night.
Some species, such as the Rock rattkesnake (Crotalus lepidus), are mainly rupicolous, others, on the contrary, called sabulicolous, mainly frequent the sandy zones.

Crotalus cerastes, here hidden in an Opuntia cladode, resembles the Saharan horned viper © G. Mazza
The most specialized is by sure the Crotalus cerastes as small rattlesnake, 50-60 cm long, resembling in an astonishing way to the Sahara horned viper, with a much similar behaviour, and which, like that one, moves by lateral undulations.
This is an interesting phenomenon of functional “convergent evolution”, which may be met also in some species of the genus Bitis of south-western Africa.
The Rattlesnakes, equipped with big cephalic plates (i.e. the genus Sistrurus), have only three species.
The biggest one, the Sistrurus catenatus, lives in the great plains of the centre of USA, from south of Nebraska to north of Mexico, with some population reaching, eastward, the Pennsylvania, where these animals are located, however, lonely in small zones of the prairies, often swampy, which have not been colonized by the forest by the end of the glaciations of the Quaternary.
The Pigmy rattlesnake (Sistrurus miliarus) is common in the south-east of USA, from central Texas to North Carolina; the Sistrurus ravus, even smaller, in southern Mexico.

The Crotalus pricei doesn’t overcome fifty cm, but the poison is the same © Dr. Giuseppe Mazza
All these serpents of the genus Sistrurus are little dangerous due to their modest size (45-95 cm), but their venom is not less toxic than that of the Crotalus.
Their diet, very varied, includes amphibians and also insects, predated especially by the small species and by the young individuals.
Also the rattleless crotalines include species with big cephalic plates as well as species with the head covered by small scales.
Until recently, most of these last were united in two different genera: the Bothrops of central and southern America, and the Trimeresurus of eastern Asia and Indonesia.
Actually, there is no morphologic or ethologic character justifying such division: in both cases, it’s matter of serpents whose overall look is similar to that of the Crotalus, but the absence of the rattle. The biggest ones, always terrestrial, may overcome the 2 m of length.
A good number of the several species under the 1-1,20 m, is of arboreal habits. Apart the fact that they have decidedly less keeled scales, these reptiles are much similar to the African genus Atheris.

Pigmy rattlesnake (Sistrurus miliarius) lives in USA, from central Texas to North Carolina © G. Mazza
The most known of the terrestrial forms, is by sure the Trimeresurus atrox (some herpetologists prefer to call it Bothrops atrox) of central and southern America, of which a variety is the famous “fer de lance” of the Martinique Island.
Being greedy of rodents (we often forget how much they are useful in this regard), they often frequent the sugar cane plantations, and seen that they are a vigorous species, fast in hitting, the incidents to the countrymen are unluckily numerous and mortal. Brazil and nearby countries are by sure the elected homelands of the great terrestrial Bothrops.
Besides the Trimeresurus atrox, are located there also species close to the 1,50 m. Instance are the Trimeresurus jararaca and the Trimeresurus alternatus.
A similar size is held also by the most dangerous among the Asian Trimeresurus such as the Trimeresurus flavoviridis, called “habu” in the Ryuokyu and Taiwan islands.
The Trimeresurus haulbacki, just smaller, is known only in northern Burma. It is an oviparous species, and it seems that the females stay watching the place where the eggs have been laid.

Bothrops alternatus common in Brazil is more than a metre an half long © Dr. Giuseppe Mazza
The small terrestrial Bothrops are spread especially in southern America, often in the mountains. Usually they have a stocky body and a choleric temper.
The Trimeresurus nummifera, of Central America, has strongly keeled scales and is called “jumping viper”, because it hit with such energy that often it completely detaches off the ground.
Two species, the Trimeresurus lansbergii of Central America and the Trimeresurus ammodytoides of Argentina, one of the few Bothrops reaching the temperate zones, have a raised muzzle like the European horned viper.
In Asia, the small Trimeresurus are rare and are found mainly up in the mountains.
The Trimeresurus monticola, which is oviparous and does not overcome the 75 cm, lives in the mountains of the Indochina peninsula and in Sumatra.
Another unusual species of the high hills of India and of Sri Lanka, the Trimeresurus macrolepis, which shows already a certain tendency to the semi-arboreal life, has the head covered by irregular but remarkably enlarged scales.

Bothrops nigroviridis marchi or Bothriechis marchi is a pitviper of Central America © Dr. Giuseppe Mazza
The Trimeresurus purpureomaculatus seems to be the only Asian member of this genus, and seems to prefer living between the rocks as a rupicolous species, near the seaside and in the mountains.
Contrary to the terrestrial forms, the arboreal ones are much more numerous in Asia than in America.
Only 8 species are known in the New World, which is less than one quarter of the Bothrops, whilst the herpetological zoologists have counted at least 20 of them in Asia, which represents more than the four fifths of the species of the region.
These arboreal crotalines are medium or small serpents (the greatest reaches a maximum of 1,20 m of length), often green, with a thin white line on the sides and a red or orange prehensile tail. There are some of them variously spotted of brown or black, typical colours of the bark of the trees and not of the leaves.
In the whole, they are ophidians less nimble than the members of the African genus Atheris and more prone to go around, occasionally, on the ground during the night. On the contrary, many stay on the bushes or the small leafy and not too tall trees, with the prehensile tail rounded on a branch.
Their alimentation, often quite varied, includes small mammals, birds, saurians and amphibians.

A Bothrops schlegeli or Bothriechis schlegelii or Trimeresurus schlegelii, decidedly disguised © G. Mazza
As a general rule, they are little aggressive animals, but as many arboreal, is we try to catch them they hit repeatedly.
Formerly, they were united by the taxonomic biologists under the name of Trimeresurus gramineus, that is “the green vipers of the trees”, known also with the name of “bamboo vipers”, extremely common all over the Asian south-east and Indonesia.
Presently, this “super species” is subdivided in numerous species, often difficult to be exactly classified and determined. The green leaf colour of the skin and the golden yellow eyes, render them, perhaps, the most beautiful vipers on the Earth.
The Trimeresurus wagleri of Malaysia, Indonesia and Philippines, spotted of black, green and yellow, is a lazy animal and much less short tempered than the previous ones and is easily found among the Mangroves.
The Trimeresurus cantori of the Nicobar and Andaman Islands (these islands form as territory of India – Union Territory – two archipelagos of the Indian Ocean placed in the Gulf of Bengal and paradoxically closer to the Burmese and Sumatran coasts than to the Indian ones), is by sure the greatest among the arboreal species of this genus, with females which may reach the length of 1,15 m. Like all the members of this genus, the males are significantly smaller.

The Trimeresurus stejnegeri is common in south eastern Asia, from India to Taiwan © Dr. Giuseppe Mazza
The arboreal species of the New World are located in Central America and in the north-eastern part of South America.
One of these, the Trimeresurus schlegeli, has small horns over the eyes.
On the whole, they are not very abundant animals, and little is known about their biology, in particular of their habits and behaviour.
We have already said that the Asian Trimeresurus have tendencies distinctly more arboreal than their American congeners, but have also another particularity: their venom is poorly toxic.
But the bites inflicted by the Trimeresurus flavoviridis, the dangerous Ryukyu habu, their bites are usually considered as benign and in any case not fatal. Strangely enough, here it realizes, biologically speaking, the opposite of what happens between the Agkistrodon, where the Asian species are much more feared than the American ones.
The last of the rattleless snakes and with the head covered by scales, is the famous South American Lachesis mutus the only member of its genus.
The length of the greatest specimens may amply overcome the 3 m, and being animals with a robust body, it may happen that their weight overcomes that of the big king cobras.

Also Trimeresurus purpureomaculatus has a vast distribution, from India to Indonesia © Dr. Giuseppe Mazza
It is a strictly terrestrial serpent, not aggressive, but fearless, whose universally accepted name of “terror of the woods” does not need any comment.
The Lachesis mutus is oviparous, and is perhaps the only American crotaline having such particularity.
All the rattleless corallines, and with wide cephalic plates, are united in the genus Agkistrodon or Ancistrodon, a more euphonic term, this last, preferred by the French biologists.
There are three species of them in America and they are commonly known as “moccasins”. The greatest, the Agkistrodon piscivorus or Ancistrodon piscivorus of the south-eastern USA, reaches the length of 1,50 m. It is a dark serpent, with much heavy body, whose semi-aquatic habits are unique among the Viperids (Viperidae). Its diet is based essentially on fishes and amphibians.
The Copperhead moccasin (Agkistrodon contortrix ), smaller, very common in the eastern part of USA, gets its name from the typical copper nuance (often with pale pink and salmon rings), present on its head.
The Mexican Agkistrodon bilineatus also terrestrial,is quite similar, but has, especially on the head, some thin very marked white stripes, which have a a very nice effect.

The Copperhead (Agkistrodon contortrix) is common in the eastern part of USA © Dr. Giuseppe Mazza
These three serpents are little dangerous, and tend to scare the opponent rather than to bite it.
The Agkistrodon piscivorus has also the habit, very unusual in the venomous species, to how open their great mouth, where the very pale mucosae stand out against the dark colour of the body.
In any case, their venom is little toxic.
Contrary to the Trimeresurus, all Asian Ancistrodon are terrestrial and are more numerous and more dangerous than the American members of the genus.
The Ancistrodon halys, spread from Japan to southern Volga and from southern Siberia to Tibet up to southern China and Taiwan, is the only crotaline reaching Europe.
Few reptiles have a repartition area so vast, and in fact this small 60 cm serpent, even if typical of the steppe, lives also in different biotopes and climates, thus denoting a very high capability of adaptation and acclimatization.
There have been described eight races or subspecies, which clearly present, besides morphological differences, also physiological ones.

If menaced, the Agkistrodon piscivorus shows, as warning, the clear buccal mucosa © G. Mazza
The Ancistrodon acutus of southern China, Taiwan and north Vietnam, reaches the 1,80 m of length, and is without any doubt the greatest member of the genus. Lazy but short-tempered, it mainly lives in the stony mountain zones covered by woods.
The Ancistrodon rhodostoma (we have not to forget that Ancistrodon is equivalent to Agkistrodon, it’s only matter of a different phonetic pronunciation), smaller but more lively, replaces the previous species in the Indo-Malayan peninsula, in Sumatra and Java.
It is the cause of several incidents, in particular while ploughing the soil for preparing the rubber plantations.
All other Asian ancistrodontes are less than 60 cm long, and the Ancistrodon nepa of Sri Lanka, does not even reach the 40 cm. They mainly eat small rodents and saurians.
The Ancistrodon himalayanus, of western Himalaya is by sure one of the serpents which reach the highest altitudes. It normally lives between the 2000 and the 3500 m, but some specimens have been seized at more than 5000 m of altitude!

The Mexican Agkistrodon bilineatus is easily recognized for the white head bands © Dr. Giuseppe Mazza
The other two species of Asian ancistrodontes, the Ancistrodon hypnale and the Ancistrodon strauchi live respectively in the south of India, with Sri Lanka, and in the mountains of south-western China.
The Ancistrodon rhodostoma is oviparous, and it seems that also in this species the females watch the place of the egg-laying and incubation of the same. This could simply mean, however, biologically speaking, that they are very sedentary animals. It is evident that in Asia the crotalines, like those of genus Agkistrodon, especially in the temperate zone or in the mountains, and the Trimeresurus, solely in the tropical and subtropical zones, have a distribution practically complementary to that of the viperines. Only the Russell’s viper (Vipera russelli) cohabits abundantly with them, even if occupying a different ecological niche, seen that in its repartition area there are no crotalines of a similar size.
At the limits of its geographical distribution, the Agkistrodon halys often comes in contact with various vipers such as the Vipera berus north, the Vipera ursinii renardi west, and the Vipera lebetina south-west, but it seems that locally there is not a real cohabitation, as the Agkistrodon halys needs of warmer and drier biotopes than the two first species of vipers, and cooler than the last one.
Comparative physiology of the organs and systems in the Ophidians

Scales of Bitis gabonica. Their number and shape are used for the systematics of the species © Dr. Giuseppe Mazza
The skin
A distinctive feature of the skin of the reptilians is that of being dry and scaly, very poor of glands.
Likely all other vertebrates, if is formed by two main layers: the epidermis, the outer layer, derived from the ectoderm, the outer germ layer of the embryo, and the derma, the most inner layer of mesodermic origin, the intermediate germ layer of the three germs, ectoderm, mesoderm and endoderm in which the embryos of the vertebrates subdivide during the ontogenetic development. The scales, characteristic common to all the reptiles belonging to the order of the Squamata, come mainly from epidermis; however, also the mesoderm takes part in their formation. In each species, the scales have a typical shape and a rather constant and specific disposition. And seen that quite often they are easy to describe and count, they furnish the taxonomists very useful classification criteria, especially when we have to make distinctions between species, subspecies or races very similar. The study of their morphology, and the counting of the rows of scales, is therefore aspects which are essential for the evolution and optimization of the systematics of the Squamata and, in the specific, of the Ophidia.
Without forgetting that the biologists do consider the skin as a true and proper organ, which in all animal species, where it is more or less specialized, particularly in the vertebrates, the same is the matrix interfacing the outer world, the environment, with the inner world of the animal.

The drawing of the skin has an important role in the camouflage of this Bitis arietans © Dr. Giuseppe Mazza
And are attributable to it, sensorial-nervous, exocrine-endocrine and ecoethologic-ecophysiologic mechanisms. For instance, the various colourations which characterize it, have essential functions in the sexual selection and therefore in the reproductive biology of a species, acting as “sexual attractors”, like the characters of sexual dimorphism, permanent or seasonal, but are useful also for the mechanisms of camouflage, disguise or concealment, for escaping from a predator or for seizing a prey.
This applies, as already briefly said, both for the mechanism of the “somatolysis” or also for the “homochromy”, as well as, in the case of the sea snakes, of the “pigmentary polarity”. And this applies for all the animal species forming the animal Kingdom and, by means of other structures, also for the members of the vegetal Kingdom.
In regard to the various coral snakes it is worthy to say briefly that during the fifties a theory was proposed about the camouflaging role of their livery, called theory of the “Mertensian mimicry”, of a rather complicated interpretation and nowadays abandoned by the biologists.
The Mertensian mimicry was discovered for the first time by the biologist W. Wickler who suggested this name in honour to R. Mertens, herpetological biologist who had studied in depth the coral snakes of the New World, animals in which this camouflaging phenomenon has been frequently noted.
The name “coral snakes” given to these reptiles, comes from the showy colouration formed by alternated bands of red, black and yellow colours, which characterizes their livery.

This splendid coral snake (Micrurus fulvius fulvius) has a deadly bite © Dr. Giuseppe Mazza
These ophidians belong to two different families, the Elapids, venomous and often mortal, and the Colubrids, harmless.
In particular, the Mertensian mimicry theory is based on three or more components with similar colouration.
In the simplest situation, there are two mimes and only model and precisely: a) a desirable colubrid (mime); b) a mortal elapid (mime) and, c), a moderately venomous elapid (model).
The species “b” even if protected, cannot represent the model, as its predator cannot be able to experience on it, as usually it dies after having attacked the prey.
In the same way, also the species “a” as harmless should not have anything to “teach” the predator in order to be avoided.
Therefore, it is the species “c”, moderately venomous, to hold the best conditions for representing the model for both mimes.
Going back to the serpents and their skin, in most of the species the lower surface is covered by only one great row of scales, called ventral, which hold an important function in the locomotion.
The presence of these great scales, which form an indispensable source of friction for creeping, characterizes the serpents, even if at times they have modified a little their role, like in the boids, with lateral ventral scales, which cover only the 2/3 of the ventral surface, in some hypogeous species belonging to the orders Typhlops and Cylindrophis, and in the sea snakes and some fresh water snakes, where these scales have become small, just little bigger than the caudal and dorsal ones.

And this harmless Lampropeltis triangulum elapsoides has strong interest in resembling to it © G. Mazza
The skin of every animal must have the possibility to renew during its life, for compensating the wear and possible lacerations.
In the mammals, man included, the dead corneous layer of the skin is continuously eliminated in tin scales, and replaced by the growth and by the keratinisation of the live deepest cells.
This method, called of “small scales continuous elimination”, seems to be utilized in the reptilians by the turtles, with loss of outer horny zones of the carapace, under form of scales, and in the crocodiles, with irregular loss of cutaneous scales in various parts of the body, in independent regimen, not synchronized. The saurians and the serpents have a different modality: these animals have, in fact, the so-called “moulting periods”.
With this mechanism, in well-defined periods, the whole superficial layer of the skin (attention, only the superficial one!) is eliminated at the same time. There is, clearly, in evolutionary terms, a certain parallelism with the moulting of some birds and of some fur animals.
Though the serpents usually eliminate their skin in its entirety, or at least in three great fragments, at times they do this in stages. It is possible that this phenomenon is due either to bad health conditions or to an unfavourable local situation, such as, for instance, the lack of humidity.
In the sea snakes, the moulting happens normally as well as by means of the elimination of the detached pieces. After two or three weeks before the moulting, the skin of the serpent saddens, its eyes become dull and bluish, possibly due to modifications of the inner structure covering the eye.

Malpolon monspessulanus exuvia. Periodically serpents renew their skin by moulting © Dr. Giuseppe Mazza
Often the animal (this is well known by the biologists which keep serpents in the terraria-vivaria inside zoological gardens and aquatic parks), refuses the food and becomes cranky, may be because of the fact that it does no longer see well.
Often, it spends more time in the water, and as we know that the permeability of the skin increases before the moulting, it is possible that in this way the serpent impregnates well its body, in order to reduce then the risk of desiccation. It may well be that the water serves to soften the keratin.
Few days before the real moulting, the membrane covering the eye becomes again transparent, and the skin assumes an almost normal colouration; we might think that the moulting has already happened, if it not was for the fact that we do not find around the cast-off skin, called “exuvia”.
The serpent is often nervous. It goes around without stopping, here and there, rubbing the head against the obstacles trying to break the skin in the zone around the mouth. The serpents as well as the saurians, have a physiologic mechanism for facilitating this process: the backflow of the venous blood from the head to the heart is in fact hampered by the constriction of the jugular veins.

In rattlesnakes the sound is produced by grits shaken in the empty corneous segments of the tail © Mazza
By that time, the head swells of blood, especially around the eyes, and facilitates the breakage of the dead corneous layer.
Once the old skin has detached around the jaws, the elimination of the whole is usually much easier. If then it gets caught in a stone or in some branches, the serpent is able to get rid of it quickly and it may happen that the skin gets turned inside out, like a sock, that is, the inside becomes outside and vice versa. Of course, as said, this process interests only the most superficial layers of the skin; the exuvia is therefore thin and frail, with a completely different structure from that of the thick skin of a serpent, and has only few traces of pigmentation. In the countryside, at times, we find the complete remains, even the membrane which surrounded the eye, and it is by means of these that the biologist is able to identify the species living in that particular zone.
The frequency of the moulting varies very much from one species to another, and also between individuals of a same species, and even for one same specimen.
The popular belief (work, alas, of many naturalists and veterinarians!) after which the reptilians have only one annual moulting in spring, does not correspond, then, to the reality. The zoological biologist Klauber, during the seventies of the last century, after having analyzed the data of many observations on the field, has concluded that in the wild the rattlesnakes moult from one to three times a year, and that the moulting is more frequent in the warm climates species, than the northern ones, and that they have only 6-8 months of annual activity.

Young Chondropython viridis with a showy spotted livery © Dr. Giuseppe Mazza
The rattlesnakes, stabulated in terraria-vivaria (and this is also confirmed by my own experience), may moult even seven times per year. And an even higher frequency has been reported for the pythons. The first moulting takes place a few days after the birth, and the biologists have seen that in some viviparous species it may happen even inside the maternal uterus!
Although much has been written about the moulting of the snakes, it’s only during the last thirty years that more detailed studies have been done about the physiology of this process, in particular on the cellular modifications which are the responsible of the same.
From deep studies of 1980 on daily samples of skin done by the biologist P.F.A. Maderson, on the Beauty rat snake (Elaphe taeniura) kept in the State aquatic park of Singapore, it has been observed that the skin of a serpent, generally speaking, enters a quiescent phase (functional rest), where we can see the simple structure, with external corneal zone, provided with grooves called “Oberhautchen” (German word which literally means: carved, the name has been kept as such, because these structures were discovered during the first half of the ‘900, by the herpetological biologists of the Zoology School of Berlin), with keratinized laminae, followed by a deep layer of basal cells, and, between the two, a double layer of more or less flattened cells. The mechanisms originating the moulting are extremely complicated.
Complicated biochemical channels activate and it is necessary that a metabolic change of the animal happens involving also cells of quite different stipetes, for instance also of immunological nature, like the eosinophil granulocytes, which from the blood reach the most external layers of the skin, and express and secrete in turn proteolytic enzymes. And these will start to digest the proteins of the epidermis, thus starting a phase of cleavage of the tissue.
These are advanced subjects of the cellular biology and of the biochemistry, which are beyond my training as well as the context of this opus, but in quite generic terms the description just done is sufficient. The zoologists have always given much weight and dedicated many studies to the colours of the animals, as the botanists do for the flowers, because, being biologists, they have immediately perceived the intimate essence hiding behind a coloured livery of an animal or the corolla of a flower, the “communication” between conspecifics and congeners and also between members of other species and genera.

But this adult Chondropython viridis has adapted the livery to its new lifestyle © Dr. Giuseppe Mazza
The colour of the skin and of the other contiguous membranes, as well as the mucosa covering the mouth and the cavities of the body, depends in large extent from the presence of particular cells containing pigment, called chromatophores.
They are placed almost always in the outer layers of the dermis, not of the epidermis, and in fact the exuviae are paler.
The most common type of cell is that containing a dark brown pigment, the “melanin”. This melanophore cell has several ramified diverticles projecting into the basal layer of the epidermis. If the cell contracts, it releases little melanin, if it relaxes expanding, the melanin will be more abundant.
This mechanism is utilized by many saurians, such as the chamaeleontides in order to vary the colour (more or less dark) of their livery, corresponding to their emotional conditions. These transitory modifications with mimetic purposes or for camouflaging, or in relation with the emotional state of the animal, are absent in the serpents, and the melanophores determine only the dark colouring of the body. The absence of melanin is the main cause of the white or very pale colour in some serpents, as we may see in the wonderful white python quoted in the book written by the biologist C.H. Pope, “The giant snakes”, of 1958.
Some biologists, during the years ’70 of the XX century, in Asia, have also found a spotted python, with great white dots. We have not to mistake, as often happens, the melanophores, cells forming a compact, subepidermal, continuous layer, with the melanocytes, cells with always melaninic contents, but located in the epidermis itself and which are those originating the very rare and very weak pigmentations present in some exuviae, seen occasionally, because dragged during the moulting.

Some serpents, like this Vipera ammodytes, show some horns © Dr. Giuseppe Mazza
However, as we have seen before in the classification of the serpents, there are quite numerous species having all the body or parts of it brightly coloured.
These colours, like the red, blue, yellow, black, brown, copper, etc., are made by pigment cells, which have been given the most various names, but which, generally, are called lipophores (because the contents is lipidic) and allophore: the nomenclature of the coloured cells, called chromocytes is complex and often confused, in this respect, a useful guide is the “Dictionary of Herpetology” written by the herpetological biologist J.A. Peters, published in 1969, still of remarkable value. We conclude the discussion about the skin treating the structures and special appendices found in the serpents. Some colubrids of the genus Natrix, and other similar genera, have, as already said briefly, under the scales, a double glandular chain at times extending for the whole length of the body.
Other genera have the same on the sides of the mid-dorsal line of the neck. Their function is not yet entirely clear; we think that they express a repellent secretion and irritant for the mucosae of the predators (also another ophidian), which then would be compelled to release the grip. And many male serpents, before mating, rub their chin on the neck and the back of the partner, probably as mechanism of courting mediated by pheromones released by these glands.
Some viperids, like the Saharan horned viper (Cerastes cerastes ), the viper Bitis cornuta and the Horned rattlesnake (Crotalus cerastes) of North America, have, under each eye, a sharp tubercle, similar to a horn, formed by an enlarged scale. The great Gaboon viper (Bitis gabonica) and the Rhinoceros viper (Bitis nasicornis) have usually a muzzle decorated with a pair of protruding tubercles, but, at times, these appendices are just visible.

Those of the Erpeton tentaculatum are even mobile © Dr. Giuseppe Mazza
Some arboreal serpents have the muzzle ending in a long point. In the colubrids of the genus Langaha of Madagascar, this appendix is quite odd, and it differs in the two sexes. For instance, in the male of the Langaha allaudi it is shaped like an elongated prominent horn, before the nostrils and at the extremity of the maxillae, whilst in the female the structure is much more complicated, assuming the shape of a leaf with jagged and fringed edges. The function of these appendices is not known, maybe they serve to better camouflage the animals in the dense tropical vegetation.
We mention one last case representing a unique example: the muzzle of an unusual Indochinese aquatic serpent, the Erpeton tentaculatum, which has two long ornamental tentacles, formed by dermal connective tissue and covered by scales, provided with mobility, but, it seems, insensitive. When the serpent is fully immerged in the water of a river or in the sea, they protrude straight forward, and then turn backward when the head emerges. After some biologists, when in water they should simulate small twigs or floating algae, useful for camouflaging and approaching easily a prey, or for defending, thanks to the camouflage, from caimans, gavials and some carnivorous marine fishes.
As briefly said, have had, evolutionary, a total regression of the pelvic and scapular girdle, being limbless. But in the Boidae we may see them at ventral level, in correspondence of what would be the sacrolumbar structure of a tetrapod vertebrate (pelvic girdle), hints of even vestigial organs, of small bony protuberances, as evolution trace of limbs now completely atrophied. Lastly, without any doubt, the most original appendix found in the splendid world of the serpents is the “rattle” of the rattlesnakes (Crotalinae).
It is, essentially, matter of a certain number of corneous segments, stuck into each other, which replace the conical caudal scale, present in all rattleless serpents. Each segment is usually trilobed; the disposition of the segments is as such that the two back lobes one segment fit within the two fore lobes of the following, but the last segment which obviously is free and therefore visible in its entirety. The sound is generated by small stones which remain trapped during its formation, in the development of the animal.
Nervous system and psychology of the serpents

Vipera raddei. The skull and so the brain of the serpents is quite small compared to the body © Dr. Giuseppe Mazza
The brain of the serpents is small if compared with that of the mammals. Often it is not bigger, at times even smaller, than the brain of an amphibian or a fish which may have a similar cranial volume.
However, when we compare the cerebral volumes, we have to keep in mind that the great sized animals tend to have a brain which, compared to the volume of the body, is comparatively smaller than that of species of minor size. It is important not to ignore this aspect of the encephalization index, when confronting the mass of the brain of animals belonging to different classes of vertebrates. The human brain, one of the largest in absolute value between the mammals, is about 1/45 (the 2,2%) of the weight of the body, whilst the tiny brain of the shrew mouse, is approximately equivalent to 1/23 (the 4,3%) of the weight of the entire animal; its “relative weight” is therefore about the double than in the man, what changes in favour of the humans is, without any doubt, the absolute weight.
For this reason, we must not be surprised that the brains of the dinosaurs were so small, in respect to their huge body size, even if in absolute value they may have been even greater of the brains of the extant reptilians. The brain of most reptilians never fills up completely the cranium, from whose walls it is separated by means of the “dura mater”, the robust external membrane wrapping it. And this, having only the skull available, renders difficult the work of the paleontological biologists which wish to reconstruct the fossil brains. For instance, in the Tyrannosaurus rex, the endocranial imprint is 20 cm long and does not exceed the 5 cm of width. Obviously, the actual measurement of the volume of the brain may be quite less, but in this way we may get at least an indication, even if approximate, of the possible absolute dimensions.

Vipera aspis. The brain of serpents is smooth, without convolutions © Dr. Giuseppe Mazza
The brain of the reptilians, and therefore of the serpents, like usually of all the vertebrates, is subdivided in three main parts: fore, mid and hind, from where depart one or more of the twelve (at times only eleven) pairs of cranial nerves. The surface of the hemispheres is usually smooth, even if at times, there may be one or two small roughness. Are therefore lacking the typical deep circumvolutions, mainly present in the human being, in the non-human primates and in the great apes as well also in the delphinids.
Most of the forebrain is formed by two symmetrical swollen masses, the cerebral hemispheres, which contain cavities called lateral ventricles. The rostral (fore) part of each hemisphere narrows into a short stem, which then swells to form the olfactory bulb, pair organ. These bulbs are in any case located just behind the nose of the serpent, and the fibres of the olfactory nerves head in the nasal pits.
In some species of serpents there is a further type of bulge, the auxiliary olfactory bulb, placed behind the main organ where go the fibres of the “vomeronasal nerve” coming from the Jacobson’s organ. In the Boidae is present this additional olfactory bulb, which adds to the two already present. A similar structure is found also in other reptilians, see Sauria and Chelonia.
As is now well known, in the human being and in the great apes, as well a in the non-human primates and the delphinids, all the cognitive-emotional sensorial processes are related and integrated at level of the cerebral cortex, called “Neo-cortex” in the human being, or commonly “grey matter”.
Many sensorial fibres reach this structure, and many muscular fibres depart from it, and go to innervate the skeleton striated muscles of the limbs and the trunk.
The aforementioned circumvolutions and the thickness (of a few tenths of mm) of the cerebral cortex, vary among the before indicated species, where, for what the thickness of the cortex is concerned, they reach maximum values in the human being, as they represent the keystones for a psychic development and a more sophisticated psychology, that is, what has, evolutionally, permitted to create those faculties, defined as “superior”, such as the learning, the memory, to reach, in humans, the verbal and abstract skills.

A charmer with a Naja naja naja, which almost surely doesn’t perceive the music © Dr. Giuseppe Mazza
In reality the brain of the reptilians, in the specific, of the ophidians, or better, the hemispheres that compose it, are mainly in relation with the smelling, as it happens in most of the so-called inferior vertebrates, definition I am not fond of. A small zone, on the upper surface of each hemisphere, is often considered homologous of the cerebral cortex; but this is a simple hypothesis advanced by the zoological biologists, not yet substantiated by the comparative neurobiologists. In terms of comparative psychology in the serpents, the experimental ablation of this zone (also for the saurians) seems to have poor influence on their behaviour, and it does not seem that there is the suppression the conditioned reflexes, learnt previously and concerning the association of the colours or of geometrical forms to the food supply. Actually, this field ought to be investigated by other types of biologists (the comparative biologists), as their approach, by means, for instance, of the electrosurgical stimulation of these areas, would be more effectual than that applied by us, zoological biologists, more based on the observation in nature or, in case, in a zoological garden, utilizing non invasive methods, which are however only able to give qualitative and not quantitative answers to the problem. Scientific studies on the comparative psychology in the serpents, through psychometric tests, are still very few in literature.
Usually, a comparative neurobiologist utilizes tortoises for his studies, whereby only the data obtained from ethologic studies remain, by observations in the wild or in controlled habitat. Therefore, to explain phenomena, such as that of the fakirs (characters I believe at least ambiguous), who would pretend to enchant even dangerous serpents, like the Asian king cobras, with the melody of the music, do not have yet any evidence as support, and therefore remain, even if fascinating, phenomena confined within the ethnology folklore. And we have to add that these gentlemen utilize always animals which have been doped or mutilated of their venom fangs, in order to render them harmless.

Thanks to the tongue and Jacobson’s organ this Boiga irregularis tastes the air © Dr. Giuseppe Mazza
Recent neuroanatomical studies, done on saurians and serpents, have shown that the ducts of the Jacobson’s organs are placed in correspondence of tip of the tongue, and the neurobiologists say that they might have a crucial role in some behaviour of these animals. The Jacobson’s organ is a pair organ (that is in double copy) in the serpents as well as in the saurians, is of spherical shape and of great size (in respect to the cerebral hemispheres), contained in a cartilaginous covering.
Also in the sea snakes, it has a remarkable size, like in most of the saurians, whilst in some Agamids (Agamidae) and in some Iguanids (Iguanidae), such as the Anolis, it has reduced dimensions or is absent, whilst in other species, such as in the Camaleontidae, it has a rudimental structure.
Indeed, for what the specialized arboreal serpents are concerned, the nose, for what I know, has never been studied and it would be interesting to do this for seeing if there are functional coincidences with the other serpents. Furthermore, in the serpents as wall as in the saurians, the organ of Jacobson is connected with determined cerebral areas of the “midbrain”, and this would explain how come that a serpent or a monitor (as example of saurians) tends to continuously evert the tongue, moving it in the air, possibly for tasting the same. What is the meaning? In a splendid documentary film of the seventies of last century, made in India and on the Komodo Island under the supervision of the great herpetological biologist Angus Bellairs of London, it was observed that pythons and monitors, when in presence of carcasses of dead animals, hidden by these scholars near their dens, suddenly did wake up from their apparent calm. They started to quickly agitate their bifid tongue, everting it, in order to localize the direction of such smell, and quickly moved, getting out from their dens on foot or crawling. This has led the zoological biologists, authors of the documentary, of the group to think that such animals, by tasting the air collect by means of the tongue small odour particles, which, through the aforementioned ducts, are conveyed to the organ of Jacobson, that, through the nasal vomer sends them in the olfactory cortex (enthorinal), where then will be probably converted (by means of discharges of electrical impulses of a specific form) in a sort of picture of the smell, by means of which the monitor as well as the python, may find their source, that is the dead carcasses.

The eyesight of serpents is modest, but some species, like Ahaetulla prasina, has a binocular vision © G. Mazza
Ear and Eye
Before, we have been talking about the smelling and the chemical organ allowing the description and the discrimination of the odours, the organ of Jacobson.
For what the other sensorial organs are concerned, such as the ear and the eye, we can say that for the first one it is difficult to determine exactly how much the serpents are capable to “hear”.
The absence of eardrum and the anatomy of the middle ear of the serpents and of the burrowing saurians, do not show by sure a great sensitiveness to the sounds transmitted through the air, even if the cochlear canal and the basilar papilla of the inner ear are not degenerated at all.
These animals have never shown in the experiments made, clear reactions to the sounds coming from the air; for this reason the zoological biologists are much sceptical about the accounts of serpents fascinated or attracted out from the dens with the music.
Nevertheless, the comparative neurobiologists Wever and Vernon, have proved that the middle ear of several colubers (genera Pituophis, Thamnophis and Natrix) respond electrically to the aerial sounds of low intensity with frequencies included between the 100 and 500 cycles/second. The problem is complicated and is still all to be resolved. For what the eye of the serpents is concerned, these pages should be just sufficient to list its complexity.

The serpents pupil can be round, slit, or locking as in this Thelotornis kirtlandii © Dr. Giuseppe Mazza
The zoological biologists Gath Underwood and Wells have performed evolutionary studies on the eyes of the serpents for about 50 years (from the thirties up to the eighties of the last century), and have been probably the greatest experts in the world. After these authors, the Ophidia, meant as group, should come from progenitors with hypogean life, whose eyes had reached an advanced stage of degeneration.
Going back to the epigean life, we think to the primitive burrowing serpents of genus Leptotyphlops and Typhlops, there have been recovery processes which have permitted them to organize the eye in evolutionary terms, in way to guarantee the life in the arboreal sub-habitat, for nocturnal life species as well as for the diurnal, arboreal, terrestrial or aquatic.
The only structure of the hypogean life still extant in the serpents as ancestral vestige, are the “spectacles” typical character of the serpents which carried life in that environment and therefore trace of a possible “troglobiotic” origin.
The adaptation and the adjustment of the eye in the extant serpents, have assumed the technique similar to that of the camera, utilized also by the selachians-elasmobranchii (sharks, dogfishes), which utilize the iris as driving force.

Boides and Crotalines, like this Trimeresurus, have thermal dimples to see at the infrared © G. Mazza
The small yellow lenses and the typical double cones of the diurnal serpents, represent advanced optical forms of the present serpents; finally, we have to keep in mind the presence of different morphologies of the pupil (slit, round and keyholeshaped), identify their preference to diurnal or nocturnal life.
The serpents with mainly diurnal life have a slit pupil (horizontal or vertical line) and those with nocturnal life have around one. Furthermore, the presence of grooves along the muzzle, like in the arboreal serpents of the genus Ahaetulla (called by some authors Dryophis), allow a binocular vision.
The nasal and labial thermal dimples are finally another means of “vision” of the serpents. They are present only in the Crotalinae and in the Boidae, whilst the vipers do not have them.
In the crotalins, they are placed as a depression between the eye and the nostril of the nose (one on each side of the muzzle), in the boas are placed at labial level.
Such structures allow perceiving the heat carried by the blood in the living organisms. It is well known, by means of the optical techniques of the infra-reds, utilized by the humans, that a warm body has a different, “red”, colouration in respect ot a cold one, which usually has a blue colour. The emission of heat, being a form of energy, has its own, well defined, electro-magnetic frequency, which may be perceived in absence of light. And then, especially the night, these structures allow these serpents to go hunting and to avoid the predators.

Fight between males or mating preliminaries of Hierophis viridiflavus © Dr. Giuseppe Mazza
Sex and reproduction
In the animal world is extraordinarily rich in imagination and at times tragic. In fact, although it has evolved to benefit of the species, it may be fatal for the single individual. When at its peak, it gets the upper hand over almost all other vital events and of the behaviour; an impressive example is the life of the Green see turtle (Chelonia mydas), which abandons habitats rich of food, performing thousands of kilometres long voyages for going to mate and lay the eggs on far away beaches, where often finds itself at the mercy of the hunger and of unpredictable dangers. The first act of the sexual reproduction is represented by the research of a suitable partner.
For us, humans, the recognition of the sexes is not a problem, but we may ask ourselves how the animals not provided of marked sexual characters may obtain this information.
Many animals solve the problem without any doubt reacting instinctively to minor somatic differences: of smell, colour and behaviour, but often, in order to facilitate the things (through the emerging and the magnifying of their structures) of the secondary sexual characters.
The French biologist Jean B.C. Lamarck and the English Charles Darwin (even although best known, it is good to remind that this last was born hundred years after the French, and has drawn many inspirations from his works), were among the first to put themselves this question: the transalpine biologist with his magnificent opus “Philosophie Zoologique” of the XVIII century, where for the first time you move from a fixist theory of the species (proposed by Buffon) to a transformative and adaptative one, so hypothesyzing first (not Darwin) the concept of “evolution”, whilst the British biologist, by the second half of the XIX century, published the opus “The descent of Man, and Selection in Relation to Sex”.
But the greatest scholars of the world of the ophidians, the first to carry on serious studies on the reproductive biology in the years ’50-’60 of the XX century, were the biologists C.M. Bogert and V.D. Roth, respectively director of the history museum and of the “reptilians department” of the zoological garden of New York.
The existence of a true and real territorialism in the serpents has not yet been completely proven, but we know that in some species the individuals regularly move around in a certain area, where they live for long periods. Furthermore, the males of many species, like the rattlesnakes, the ancistrodontes, the vipers, the mambas and many colubrids, engage in ritualized struggles, without caring about reciprocal damages: winding their bodies in a characteristic way, at times they get up and push strongly, until one of the two accepts the defeat and runs off.

The Thamnophis elegans winter together. By wake they mate in group with legendary knots © G. Mazza
The males of the nice Aesculapian snake of southern Europe (Elaphe longissima), assume a lyre-shaped position, which is probably what has inspired the shape of Hermes, or Mercury, Caduceus. This, as well as the rod of Asclepius (a serpent rolled around a staff), have become then the symbol of the medical profession, even if such profession doesn’t hold any relation with the zoology.
We have to add that, often, this struggle between two Aesculapian snake males was interpreted as a form of courting, a sort of nuptial dance between male and female; and vice versa, a coupling phase between male and female, and also the struggle between two conspecific males. The males and females of this species do not have evident somatic characters of sexual dimorphism, and it is not easy to distinguish them.
Indeed, for some species, there is a certain resemblance between some phases of the courting and a struggle, as is the case, for instance, of the King cobra (Ophiophagus hannah), where the raising of the head between male and female show similitude with a possible fight. Also the vipers, like Vipera latasti and Vipera berus, have these resemblances.
In the temperate regions, and especially in the cold zones where live the serpents of the genus Thamnophis and some species of vipers and of rattlesnakes, the animals hibernate usually together in deep common dens, and when they get out, in spring, certainly do not have a hard work to do for finding a partner. At times, they group together and mate, and this fact has originated the legend of the “knots of serpents”, of which at times we hear telling even now in the countryside. These gregarious couplings are on the contrary rare in the tropical species, which do not have the winter lethargy.
The nuptial parades of the serpents where carefully described in an article of the years ’60 of the last century, written by the zoological biologist G.K. Noble, one of the pioneers of the experimental herpetology. Usually, before mating, the male follows the female, rubbing its head and neck against its body and pulling in and out the tongue. This behaviour has been noted, inter alia, in the king cobra, where the male rubs clearly its nose on the spread hood of the female.

The mating males of the Coronella austriaca catch the head of the female © Dr. Giuseppe Mazza
The twisting of the tails is also part of the courting: the male uses in fact the tail for lifting that of the female, in such a way that the two cloacal regions may get in touch.
Later on, the body of the male is run by undulations going through its whole, from the head to the tail, and which may last up to ten minutes. After some zoological biologists, these precede the copulation, rather than the actual ejaculation. In the rattlesnakes, the body of the male is shaken by sudden jerks and not by undulations; the male boas and pythons, on the contrary, use the rudimental-vestigial hind limbs for rubbing the sides of the female, thus producing a hoarse noise, which may be heard even at 1 m of distance.
This stimulus would induce the female to assume a position such that the male may slide its tail under that of the partner and penetrate it with its hemipenis. In the Smooth snake (Coronella austriaca), as in few other species, the male grasps with the jaw the head of the female, as also happens in many saurians. The copulation may last one hour or even more.
At times, in captivity (zoological gardens and aquatic parks), they have observed attempts of homosexual couplings, indices of aberrant behaviours, due probably by alterations of the haematic title of the sexual hormones, possibly casual events induced by the stress and in any case rare in the wild. The consistency of the eggs (which may be oval, ellipsoidal and usually are pale white or white-beige in the various species of serpents), is pergameneus. They have not remarked, but in the pythonids, parental cares between mothers and newborn, in the various species, as absence of brooding. In the viviparous species, it may happen that a dead mother (for instance in the vipers) has, once dissected, great developing fetuses inside the uteruses. This, in past, generated the legend after which these serpents did swallow their own cubs in order to protect them until completion of the development; this one, in reality, takes place outside, once delivered.
Now let us take a brief glance to the nature of the chromosomes in the reptilians, and to the karyotype of the serpents. Remembering that the reptilians, and therefore the serpents, are gonochoric organisms (with separate sexes), but some cases of hermaphrodism we may observe in a family of saurians, the Scincids (Scincidae), and with amphimictic or amphigonic reproduction (by means of fusion of one sperm with an oosphere), therefore, essentially, syngamic organisms, have therefore a diploid karyotype = 2N (total diploidy).

The females of the Naja naja retain the sperm for long time after the coupling © Dr. Giuseppe Mazza
And some lizards, such those of the Caucasian walls (Lacerta saxicola), have a parthenogenetic-unisexual reproduction: the oosphere in order to activate as embryo, does not need the spermatozoon to fecundate it, but its two nucleoli, perceiving adequate stimuli of biologic, physic or chemical nature, melt thus restoring the diploidy 2N.
These “parthenogenetic” strains seem to generate in geographic areas with unstable and difficult climate, as was, in fact, the Caucasus during the Pleistocene (Quaternary) glaciations. In that period, and in those environmental conditions, the “natural selection” has probably favoured a unisexual reproduction.
Maybe because the parthenogenetic females lay usually more eggs than the syngamic ones and because they have a shorter incubation period; for this reason, they are in fact called “sudden”, distinguishing them from the “lasting”, that is those produced by amphigony, where the incubation is longer. And this clearly permitted a greater possibility of survival in those so difficult geographic areas.
The reptilian chromosomes have a variable size: there are big ones (macrochromosomes), small (microchromosomes), and intermediate. When observed at the microscope, they may have the shape of a “J” or a “V” (metacentric chromosomes), or they may be similar to rods of various dimensions “I, i” (acrocentric chromosomes). During the course of the evolution, it is possible that, with the sourcing of a new species, the formula of the karyotype may have had alterations, for instance for the fusion of two rod shaped chromosomes (acrocentric) which have led to the formation of a V-shaped metacentric chromosome.

The Bitis arietans delivers 20-50 cubs each summer © Dr. Giuseppe Mazza
Since, it is possible to establish a “fundamental number: NF” of chromosomes, assigning each type of chromosome a specific numerical value, for instance: for the chromosome “V” the value is 2 (that is, 1 per each lateral arm of the V) and the value 1 per each rod chromosome “I, i” or per each punctiform chromosome indicated with “m”, typical of the serpents. From this classification of the chromosomes, the herpetological biologist R. Matthey, who has realized extensive studies on the karyotypes and on the cytogenetics of the Lepidosauria, subclass to which pertains the suborder of the Ophidia, established that the serpents are a group more homogenous than that of the Sauria, for what concerns the structure and the nature of the karyotype.
While studying the various species of serpents, related to numerous genera, including the primitive hypogean serpent Leptotyphlops phillipsi, he remarked that most had a keryotype almost identical, with the formule 2N=36, that is 2N=10 V + 6 I + 20 m, from which, remembering that V has the value 2, whilst to I and m the value 1, we obtain the fundamental number NF = 46. This is considered by the herpetological biologists the “base formula”, of the members belonging to the suborder of the Ophidia.
The coluber Clelia occipitomaculata, has the highest known number of chromosomes, not only among the ophidians, but also among all reptilians, with formula 2N = 50; of which 14 are macrochromosomes and 36 microchromosomes. As well known in Biology, in the higher vertebrates (birds and mammals), the sex is determined by heterochromosomes (sexual chromosome). In the specific, (see the Psittaciformes), in the mammals are the chromosome X = feminine chromosome and Y = masculine chromosome, in the birds, W and Z, from which the heterogamety or heterogamy, in the mammals is the masculine “XY”, that is the male is heterozygote for the sex, the homogamety or homogamy is that feminine “XX”, that is the female is homozygote for the sex, with a double dose of the sexual chromosome X, one of which is condensed or inactivated, forming the so-called Barr body, hyperchromic (thicker, if observed with the microscope).

The Trimeresurus wagleri shows a certain sexual dimorphism with males smaller of 1/4 © Mazza
Therefore, in the mammals it’s the male who determines the sex: in the mammals the ovules are provided only of the feminine chromosome X, whilst by the males are produced X as well Y spermatozoa, reminding that the germinal cells (spermatozoa and oospheres), are haploids N, therefore they can carry only one sexual chromosome.
In the birds, it’s the contrary: the heterogamy (WZ) is typical of the feminine sex, therefore the female is heterozygote for the sex, whilst the homogamy is masculine (ZZ), that is, homozygote for the sex.
It is therefore the female to determine the sex with oospheres endowed with the chromosome W or Z, whilst the sperms of the masculine birds are always homozygotic Z.
The phase WW is incompatible with the life.
Situations similar to the birds are found for the lepidopterans and several species of coleopterans, whilst in the orthopterans, as we shall see in the relevant text, the things still a bit different. In many species of colubrids, and of viperids, the story is similar to the birds.
However, in various species belonging to the Boidae, like in members of the subclass of the Archosauria (orde Crocodilia, suborder Eusuchia, family Crocodylidae), or those relating the order of the Chelonia and the suborder of the Sauria,it has not been possible to highlight chromosomic differences between the two sexes.
Probably, from the biological point of view, in many species of serpents as well as of saurians, crocodiles and chelonians, the determination of the sex depends as for many fishes from specific single genes and not from complete chromosomes (these ideas and studies, are however very complicated and the molecular biologists and the geneticists carry them on since years). Recent researches seem to indicate that, at least in some saurians, the male is heterozygote like the mammals.
Venomous apparatus
As before described, the structure observed in the species such as the Ringed snake (Natrix natrix), in which none of the teeth is specialized to the conveyance of the venom, is called “aglyphous”. We remind that such classification, even if abandoned, is useful for understanding which genera of serpents are venomous and which not, furthermore it gives us a hint about the mechanics of the bite.

A Psammophis punctulatus trivirgatus eats the frog it has just seized. Most of the colubrids have all their teeth alike and are not venomous. But some of them have on the palate a special gland, the Duvernoy’s one, with a toxic secretion meant for their small preys. When the venom is more active and this is connected to bigger poste- rior teeth, with furrows or grooved drains for directing the venom, these animals are called opistoglyphous, and they may be dangerous for the man © Dr. Giuseppe Mazza
The aglyphous structure is found in many colubrids, as well as in all the members of the primitive families belonging to the infraorders of the Henophidians (Henophidia) and of the Scolecophidians (Scolecophidia).)
In most of the aglyphous the size of the maxillary teeth is little variable.

The Boomslang (Dispholidus typus) is and African opistoglyphous, often origin of accidents © G. Mazza
But in many colubers, the upper parts of the superior labial gland may be surrounded by a real capsule. When the distinction becomes so evident, to such region is given the name of “Parotid gland” (we remind that this has nothing to do with the parotids of the mammals), called also “Duvernoy’s gland”, after the name of the biologist who individuated it in 1832.
In some colubers, the secretion of this gland is toxic and this gland is often associated, but not always, to one or two teeth, bigger than the others. Actually, most of the aglyphous colubers are not venomous; and when they are so, like the ringed snake, that is only towards amphibians or saurians of which it nourishes, so they are not dangerous for the man.
When the Duvernoy’s gland is associated to teeth placed in a rear position in the maxilla, the serpents are called “opisthoglyphous”. These teeth have a groove, or channel, which goes through, connecting them with the reserve of venom.
The venomous opisthoglyphous serpents, as seen, cover about one third of the family of the colubrids, among these we find the Masked water snake (Homalopsis buccata), the Golden flying snake of the eastern regions (Chrysopela ornata) and the Boomslang (Dispholidus typus), an African colubrid which may aver mortal for the man.
But the boomslang, however, the venomous opisthoglyphous serpents do not secrete a venom not so toxic and even their mechanics of bite is not so efficient to cause the death in a human, but may be such only for various species of reptilians, amphibians, small mammals, and birds. In any case, when they bite, they cause a considerable pain.

The elapids, like this Maticora bivirgata flaviceps, have the venomous teeth in a fore position © G. Mazza
In the Elapids (Elapidae), like the cobras, the kraits, and their hydrophiid relatives, that is the sea snakes (Hidrophiidae), the situation is different: the venom gland is completely separated from the superior labial glands and is located behind the ear.
It is usually very big and in some species, such as in the Coral serpent (genus Maticora), it extends widely on the trunk, for about one fourth of its length.
A muscle which is attached to the glandular capsule, the “adductor mandibulae superficialis”, when contracting at the time of the bite, helps in squeezing and inoculating the venom.
In the elapids and the hydrophiids these venomous teeth are placed forward in the maxilla and are defined “proteroglyphous”.
In the cobras the margins of the groove of these teeth of conveyance of the venom melt to form a real channel, which facilitates the emission and the input of the toxic: efficiency, therefore, not only of the venom, but also of the venomous bite.
Last, the Viperids, the vipers (Viperidae), which have a structure similar to that of the elapids, but the teeth are not always fully channelled, like the hypodermic needle of a syringe, and also there is an additional accessory gland, possibly of venomous nature or of strengthening.
Such serpents, which have the most efficient venom apparatus in the world of the “ohipdians”, are called “solenoglyphous”. The spitting serpents, which include some cobras such as the African Hemachatus haemachatus and the Black necked cobra (Naja nigricollis), have developed eco-physiologic adaptations thanks to which the can spray drops of venom to a short or long distance.

The Hemachatus haemachatus may blind preys and predator, spitting them its venom in the eyes © G. Mazza
These drops, entering inside the eye, for instance of a small mammal, may blind it, and the serpent will take advantage of this for swallowing it (the reptilians do not chew).
This is well known by the biologists breeding these animals inside the terrariums in the context of zoological gardens and aquatic parks. When disturbed, they can spray the venom on the glasses of the terrarium, and without protection, they might blind also the man.
Nature of the ophidian’s venom
The venom of the serpents (ophidians) has a highly complex structure from the biochemical point of view; it contains many enzymes, such as the protease, the cholinesterase, the ribonuclease and the hyaluronidase, besides non enzymatic proteinic substances.
Acting alone or combined, these substances obtain devastating effects when they are introduced in the tissues of other animals and are carried by the circulatory system and by the lymphatic system. Some non enzymatic proteins, such as the “crotamine”, obtained from the venom of the tropical rattlesnakes, stand probably among the substances responsible of the most serious damages, death included. The immunity to its own venom, by the serpent, comes from its capacity to elaborate substances (immune: antibodies as well as antagonist) antidotical, capable to neutralize it.

But it’s the Viperidae which hold the most efficient venomous apparatus, like this Bitis worthingtoni © Mazza
This autoimmunity becomes manifest only some time after the birth when the serpents matures its venom gland and therefore may be cause of mortal incidents, when conspecific adult species bite the newborn, and different races of the same species or coming form geographically different areas. Because of the biochemical venom it ejects, every genus or species of serpent, biting, causes in the prey and in the man, effects of different nature, which may be listed as follows:
–Neurotoxic effects on the brain, the spinal cord, nerve terminals, etc., neurotoxic venom.
–Effects on heart and lungs (heart attack, respiratory arrest), cardio-pneumo toxic venom.
–Alteration of blood vessels causing haemorrhages, vasotoxic venom
–Blood clotting (this and the previous effect, antagonist, may be present also in the venom of a same serpent, thus complicating its clinical situation), haemotoxic venom.
–Haemolysis (destruction of the red cells), haemotoxic venom.
–General deterioration of the tissues (necrosis and gangrene), cytotoxic venom.
Probably, in the last case, the cytotoxicity is due to an agent, the “hyaluronidase”, an enzyme inducing the intercellular destruction of the animal tissues. To complicate the picture (thus advantaging the toxicity) of poisoning, there are substances which the bitten animal and human release due to cellular and tissue reactions, such as the bradykinin and the histamine, which contribute to the loss of the physiologic balances of the organism.
Therapeutic treatment after serpent’s bite
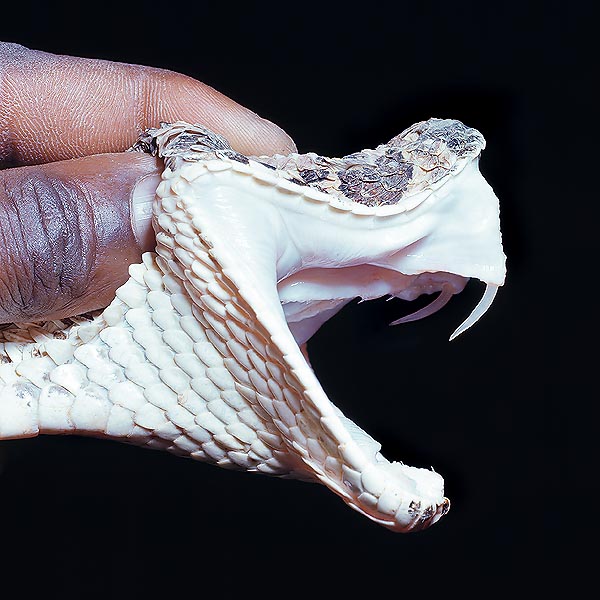
This fearful Bitis arietans, exhibits enormous teeth, folding and hollow as a syringe needle © G. Mazza
Only a professional herpetological biologist, with deep experience on the field, is capable to establish if a bite and consequent inoculation of venom have occurred, and, from the morphology of the injury, say which kind of serpent has done this, practicing the first forms of first aid and indicating which antiophidic serum is to be given.
To establish if or not the bite and therefore the inoculation of the venom has happened, is essential, as administering an antiophidic serum, in absence of the venom to fight, might cause dangerous allergic reactions, as dangerous as the venom itself, resulting in the death of the hapless getting it. For such reasons, the person subject to a bite must be immediately carried to a first aid centre (we are playing with life and death), as only a biologist or a physician may intervene.
The uninitiated, even if passionate of serpents, but not holding the technical knowledge, must avoid treatments which might only worsen the clinical case. The present knowledge and techniques of intervention have not so much evolved since the years ’60 of the last century. And this due to a general lack of interest on the part of the doctors, about the studies done by the biologists (zoologists, herpetologists). We have to say, however, that many antiophidic serums (antiviperids, antielapids: cobras and kraits, etc.) are available in specialized centres.
Institutes such as the Butantan of Sao Paulo, Brazil or the Institute Haffkine of Bombay, India, as others in USA and in Australia produce and have available antivenom serums for several species. Some of them are monovalent (that is specific for the venom of a single species of serpent), others, polyvalent (they are antidotic cocktails for numerous venomous species of serpents). Poison control centres in Italy are located in Bologna (Ospedale Maggiore), Catania (Ospedale Garibaldi), Cesena (Ospedale Maurizio Bufalini), Roma (Policlinico Agostino Gemelli), Milano (Ospedale Niguarda) and in many other cities.

Only sure remedy is to extract the venom and prepare a specific anti-ophidian serum © Dr. Giuseppe Mazza
Though there are still divergent opnions, theoretically, the first intervention (as per the F.E Russel “Cyclopedia of Medicine, Surgery and Specialties” of 1962 and, H.A Reid “Snakebite in Tropics”, British Medical Journal of 1968), should aim to the extraction of the maximum possible quantity of inoculated venom, before the same goes into circle in the organism.
This may be done binding at first, with a tourniquet (or, if we are in the country also with blades of grass or with the belt), upstream of the wound, and then doing a small cut which connects the two small of holes of entrance of the teeth (paying attention not to effect too deep cuts in order to avoid lesions to the tendon), then suck the blood, spitting it at once.
Swallowing venom of Common European viper (Vipera berus) or of the King cobra (Hamadryas hannah), as well as that of other venomous species, in reality doesn’t poison those who do so, because the venoms are effective only when in the blood system, but if the person providing assistance, sucking the blood from the injury, has some injuries in the oral cavity (for instance, an extracted tooth), or small cuts on the lips, such as to put in circulation the sucked venom, then he will poisoned too.
You must then apply a stick of wood, or an elastic band, in order to reduce at the minimum the movement of the region bitten, thus contributing to reduce even more the putting into circulation of the venom. This applies for bites received on the limbs. The matter complicates remarkably in case of other parts of the body, for instance the neck. To avoid, in all ways, the utilization of ammonia-based compounds on the injuries and give alcohol to drink (the alcohol causes vasodilation, thus increasing the possibility of putting in circle of the venom), conversely, giving water or warm and sweetened teas relieves. Lastly, the antiophidic serum must be given in an hospital structure (for this reason the subject must be at once carried in hospital), in such a way that the doctors may monitor also the possible allergic reactions which might occur, taking proper action; reactions which, as said before, are equally dangerous.
Serpents’ major diseases

The Ahaetulla nasuta, due to the odd shape of the head, is often held in captivity © Dr. Giuseppe Mazza
As briefly said before, I am not a great expert of serpents’ diseases, and this is a matter also little treated by the veterinarians, therefore we shall do a short description of these, after having done a short mention to the lymphatic and immune systems of the serpents. Little is known about the cellular and chemical defence means used by the serpents, or the reptilians in general, for protecting themselves against traumas and illnesses, but everything leads us to believe that they are basically similar to those of the other superior vertebrates. The lymphatic system of the reptilians has lines common in the class, for what concerns the ganglia scattered in the rear part of the belly, the tonsils placed in the covering of the pharynx and the islands of thymus in the region of the neck. In the serpents, as in the saurians, usually these last ones are two pairs, placed on each side of the trachea, not far away from the thyroid; they have not identified any organ analogous to the “Bursa of Fabricius”, typical of the birds. The comparative pathology of the serpents is little developed, as well as that of the other reptilians, amphibians and fishes. Among the most common diseases of bacterial origin, are the “mouth ulcers”, observed in captive specimens and seemingly due to pathogenic species such as Pseudomanas and Pasteurella, and several forms of bacilli septicaemia (Aeromonas or Proteus), and passed by mites.
Also the TB, caused by several strains of Mycobacterium, is well known. Mycoses are less frequent in the reptilians, and therefore, in the serpents, than in the amphibians, even if in the captive turtles they may at times originate sores around their eyes. Most of the infectious diseases are caused by protozoa or bigger and multicellular parasites. For instance, the Entomoeba invadens is an intestinal parasite which causes abscesses in the liver of the serpents, and also in other reptilians and in the amphibians. In the blood are found plasmodia and trypanosomes similar to those of the malaria and of the trypanosomiasis, which are transmitted by hematophagous insects (nematocera and brachycerous diptera: mosquitos and flies), whose stings penetrates through one scale and the other, or in the buccal mucosae.

Many collectors would forge papers for having this false coral snake (Lampropeltis zonata) © Mazza
The reptilians and therefore the serpents also host their part of parasitic worms, of the most different types: flat, cylindrical, hooked or with cups. There are ticks, various mites, and the workers in the reptile zoos are particularly afraid (myself included) of the infections of Ophionyssus, because these arthropods transmit infectious diseases which may quickly lead to death. Usually, in the reptilians (and therefore in the serpents), are recommended treatments with paraffin oil or with powders of the type “silicone spray”.
The most unusual reptilians parasites are maybe the odd “liguatulina”, which even if they seem worms, are in reality degenerated arachnids (spiders) which live inside the lungs (or other cavities) of the tropical serpents and other reptilians.
Only the vital cycle of very few species is known and it is possible that some of these spend their whole life in the same host. Others, on the contrary, like the Porocephalus crotali, during the larval stages are parasites of rodents and amphibians, usual preys of the reptilians and of the serpents in the specific, and become adult in various organs of the rattlesnakes. There are also species of flies (besides those transporting the Tripanosoma), which lay eggs in whatever wound present in a serpent, as well as in other reptilians and in the cloaca of the tortoises. The consequent birth of the larvae causes “myiasis”: that is, the production of aberrant cutaneous outgrowths. Other diseases are only at the beginning of their study, for instance, it has been observed that some serpents have, in artificial containing locations not enough vast, an atheromatous degeneration of the arteries, similar to that of the mammals, man included. There are both benign and malignant tumours; epithelial cancer (carcinomas), or of the pigmented tissues (melanomas), as well as tumours of bones and the cartilage. In the vivaria frequently take form keratinized warty growths, maybe caused by mites; it is thought that the tumour called “tree bark” present in the reptilians might be a form of this cutaneous degeneration.

Some, despite its dangerousness, would even keep at home a Crotalus basiliscus © Dr. Giuseppe Mazza
The reptilians, and so the serpents, do not seem to be able to catch and not even transmit the rabies, but further proofs are needed before being able to exclude them completely from this disease.
Perhaps, some epidemic forms of salmonella, which in the seventies of the XX century have been much frequent in the UK and in USA, especially on the children, might be associated with a transmission made by tortoises kept as pets.
Conclusions
Lots of people harbour a strong aversion for the reptilians, especially for the serpents, and there is the tendency to attribute this phenomenon to a deeply rooted instinct, inherited from our old ancestors.
But most of the other mammals are not at all scared by these animals: mice, rats, offered as food to captive serpents, before being attacked, run over their body with perfect irresponsibility, and it happens, at times, as observed by some biologist of the vivaria-reptile zoos, that they are themselves, the rats, to kill the serpents. A research of the second half of the seventies of the XX century has shown that the Rhesus monkeys (i.e. Macaca rhesus) bred in captivity in a zoological garden react much less intensively to the presence of the serpents than what is done by the specimens which have been seized when adult. This difference may be due to the fact that their behaviour depends, at least partly, from the environment where they have grown up, and maybe from the learning due to the direct experience or the example. Some American psychologists have proved that very young children (0,5-1 year of age) are not afraid of the serpents, but that often this fear generates before they get 4 years old.

Whatever dangerous or frail as this nice Opheodrys aestivus, it is easy to understand, also from this big list of diseases, that keeping serpents it is a specialists’ matter, admissible only for instruction or study purposes. Like ever living being they are to be respected because they have a role in the balance of the nature. The control the growth of pests and from their poison medicines are obtained © Dr. Giuseppe Mazza
It is not clear whether all this is due to the maturing of an instinct or to the example. The zoological biologists Desmond and Romina Morris, published in a book, by the end of the sixties of the XX century (Men-Animals), the results of an enquiry held on a sample of about 12.000 children, concerning their attitude towards several animals and reptilians, in the specific, serpents.
The last ones did occupy, by far, the first position in the listing of the disagreeable among the guests of the zoo of London.
However, it is a consolation to us biologists that some children (0,9% males and 0,3% females of the sample) have chosen the serpent as preferred animal.
We have seen in this text, which I deem anyway incomplete, how (and I hope this applies also to you) is fascinating the world of the serpents and how much these animals are complex.
Which is why, next time you will go in a zoological garden or an aquatic park, remember not to consider them, as often happens too quickly, among the most stupid and ugliest animals of the nature, and not to forget that without the reptilians, and therefore the serpents, ultimately, we, the humans, would not exist, evolutionary speaking !