
Text © Prof. Pietro Pavone

English translation by Mario Beltramini

Brassica oleracea var. acephala is an ornamental cauliflower. They look like a flower but are leaves, and the cauliflower we eat is not a flower but a showy reserve organ © Dr. Giuseppe Mazza
The name of the family has been proposed by the botanist Théodore Caruel (1830-1898) of French-English parentage, professor of botany and curator of the Orto Botanico (Botanical Garden) of Pisa and director of the Giardino dei Semplici (Garden of the Simples) of Florence.
The family gets its name from the genus Brassica, Latin term of the cabbage, introduced in literature by Plautus (III-II century BC), and other Authors. However, the origin of the name is uncertain and probably goes back to the Greek epithet Βράσκη (braske) or the Celtic bresic (cabbage). Yore, the family was called Cruciferae and this epithet had its origin from the late Latin crucĭfer (= cross-bearing), because of the four petals arranged in the shape of a cross.
For a long time, the Brassicaceae have been considered a natural group and Carl Linnaeus (1707-1778) himself in his “Sexual System” (Species Plantarum tome II, 1753) has included them in the fifteenth class that he called Tetradynamia due to the typology of the stamens, six in number, four of which are longer and equal to each other, and two shorter, equal to each other too (tetradynamous stamens). More recently, some Authors have included the Brassicaceae and the Capparaceae (comprising the Cleomaceae) in one only family, the Brassicaceae.

This is actually the real flower of the countless varieties of cabbages and cauliflowers: four cross-shaped petals hence the old name of Cruciferae given to the family. Conversely, the recent name of Brassicaceae honours the genus Brassica, due to its huge importance in human feeding © Dr. Giuseppe Mazza
Molecular studies have recognized these three families closely related as having one common ancestor. The Brassicaceae are, then, sister group of the Cleomaceae and both are sister group of the Capparaceae.
In their turn, the three families have been included in the order Brassicales that comprises taxa containing uniquely glucosinates (glycosidic compounds containing sulphur), cells with the myrosin enzyme, racemose inflorescences, superior ovary, petals often unguiculate, that is, having a narrow base (petals with nail) and a series of other features common to the three families.
Also the classification inside the family has not been always approved by all botanists. The one proposed by Otto Eugen Schulz (1874-1936) has been utilized for more than seventy years, though some scholars have amply proven its artificiality. In fact, Schulz had divided the family in 19 tribes and 30 subtribes, based on different characters, such as, for instance, the ratio length-width of the fruits, compression and dehiscence of them, position of the cotyledon, orientation of the sepals, etc. However, these characters have formed in all the family due to the phenomena of convergence of the action of the environmental pressures, therefore almost all morphological characters, especially of the fruits and of the embryos, that have been very utilized in delimiting genera and tribes, have evolved independently. Consequently, the systematics he proposed could not be deemed valid. As a matter of fact, the tribes and the subtribes he proposed, in the light of the molecular studies, have resulted being polyphyletic, excepting the tribe Brassiceae that, conversely, is monophyletic.
If on the one hand the morphological data alone are not always reliable for establishing the phylogenetic relations, the input given by the molecular studies on all genera of the family, the tribes and the species is still not completed and is still under study.
However, it is not certain that morphological characters like the zygomorphism, the apetaly, the reduction of the number of the stamens to four, the merging of their filaments, etc. have evolved independently. So, for a correct taxonomic classification, an attempt is made to re-examine the morphology in the light of the molecular data and this is essential to understand the function of the homoplasy and therefore of the phenomena of convergence, from that of the phylogenetic derivation as it happens, instead, in the homology.
Also the karyological studies, with the relevant chromosome numbers, are important for the taxonomy, even if they are not yet known for all the species of the family.It has been also observed that the reduction to n = 4 of chromosomes in the Brassicaceae has taken place independently as it can be noted in the Australian genus Stenopetalum (S. nutans F. Muell.) and in many North American species of Physaria.
A great advantage for the systematics of this family has been obtained with the complete sequencing of the genome of Arabidopsis thaliana (L.) Heynh. that paved the way for a better comprehension of the genetics, of the molecular and cellular biology of the vegetables.

The genome complete sequencing of the Arabidopsis thaliana, a short cycle species, now a weed in all temperate and alpine climate zones, has revealed very precious for Brassicaceae present systematics and has paved the way to a better understanding of the vegetables genetics, of molecular and cellular biology © Jean Louis Verbruggen
In this treatise we have adopted the taxonomy proposed by the German Foundation DFG (German Research Foundation) in the online database BrassiBase realized after the priority program SPP 1529 “Adaptomics – Evolutionary plant solutions to ecological challenges / Molecular mechanisms underlying adaptive traits in the Brassicaceae s.l.” (2011-2018).
Presently, the family includes 52 tribes, 340 genera and 4.636 species, but these data are not definitive as new species do add continuously and are still to be clarified the relations between some lineages of the family.
The Brassicaceae are present in the whole world, but mainly in the cooler regions of the northern hemisphere, with the highest diversity in the Iranian-Turanian region and in the Mediterranean (ca. 260 genera) and in North America (ca. 98 genera). In Australia 56 genera are present, in China 103 genera, in France 69 genera, in Japan 45 genera, in Greece 72 genera, in Iran 100 genera, in Iraq 78 genera, in Israel 68 genera, in Italy 74 genera, in Morocco 86 genera, in Mexico 61 genera, in Portugal 50 genera, in the United Kingdom and Ireland, 50 genera, in Russia 95 genera, in Spain 75 genera.

Hornungia alpina is an orophyte present on central and south Europe mountains from 1700 and 2600 m of altitude © Gertjan van Noord
Some species reach high alpine altitudes like Hornungia alpina (L.) O. Appel and others adapt to the steppe zones such as the Rose of Jericho, o Mary’s flower (Anastatica hierochuntica L.), native to Middle East, annual plant that, during the dry season contracts the branches bearing the seeds and form a spheroidal mass that the wind pushes on the sand. During the rain season the wet branches, by mechanical effect, open (hygrochasic movement) and the pouring water frees the seeds (rain hydrochoric dispersion) that germinate in a very short time.
The presence of the Brassicaceae in the phanerogamic flora decreases as we go away from the polar regions. In the island of Spitsbergen (Norway), they represent 19% of the present flora, whilst in the Republic of Senegal only the 1%. The northern hemisphere is richer in species, grouped in about 250 genera, whilst the southern hemisphere is poorer with about 100 species.
Molecular studies show that the genus Aethionema R.Br. of the tribe Aethionemeae, is the most “basal”of the family.

Aethionema thomasianum, also orophyte from 1800-2500 m of altitude, is a rare endemism of the Graian Alps in the Cogne Valley (Italy) © Dr. Giuseppe Mazza
The divergence between Aethionema and the central group of the Brassicaceae has occurred quite probably during the Eocene, almost 32 to 24 million years ago. It is probable that in the vast geographic region of Europe, Mediterranean Africa, Saharo-Arabian and/or Iranian/Turanian, has formed the most recent common ancestor of all extant Brassicaceae, when in all the world a warm and humid climate did prevail.
The genus Aethionema, present along the “Anatolian diagonal” (located at the border between Europe, Middle East, Central Asia and Africa), during the Pliocene, had a high diversification with consequent dispersion of its species, thanks to the local geologic events, such as the upheaval of the Anatolian and Iranian highlands and the formation of the main mountain ranges of the Iranian-Turanian region.
The tribe Aethionemeae represents an essential group that allows us to detect the evolution of the genome and the phylogenetic relations inside the Brassicaceae.

Some species have adapted to live in arid zones, like the famous Rose of Jericho or Mary’s flower (Anastatica hierochuntica) native to the Middle East. Annual plant, in the dry season folds the branches bearing seeds and forms a spheroid mass the wind rolls on the sand. In the rain season the wet branches open due a mechanical effect and the heavy rain frees the seeds that germinates very quickly © Jean-Claude Thiaudière – Abdelmonaim Homrani
The colonization of the newly-formed arid and semi-arid areas in all the world has been possible, most likely, also through the system of autogamous reproduction of these plants and by diaspora, that have allowed the intercontinental dispersion of mucilaginous seeds, by means of the birds to which they adhered.
A fossil fruit of the plant, attributed to the genus Thlaspi, called Thlaspi primaevum Becker, has been found in the Ruby Valley in south-western Montana (USA) and assessed to date back to late Oligocene (30,8-29,2 million years ago).
Several Authors estimate that the family Brassicaceae can have originated about 64 million years ago and that the separation from the crown group Aethionemeae, based on the molecular dating, took place about 40-42 million years ago, whilst paleobotany and paleoecology detect a Miocene radiation.
It is sure, however, that the radiation of the Brassicaceae has been mostly determined by the Miocene climatic changes (15-11 million years ago) that have produced drier and more open habitats whose niches have been occupied by various species of the family.
This has been possible due to duplication events of the entire genome (WGD: whole genome duplications) that in most cases, have been rapidly stabilized and reorganized thus to carry out an important rôle in the genetic diversification and in the radiation of the species.
It is generally believed that the polyploidy furnishes new genetic functions and extends the resilience of the lethal mutations, increases the rate of speciation, therefore an increase of species richness, besides furnishing the adaptive advantage for colonizing difficult and unstable habitats.
As a matter of fact, the polyploid genomes have defeated the diploid ones, less competitive, when new niches have appeared due to the climate changes.
Arabidopsis thaliana has provided evidence of the multiple events of polyploidization that have led to the formation of many paleopolyploids in the family Brassicaceae as well as in the order Brassicales.
The adaptive advantage of the polyploidy is nowadays well shown in the genus Draba (tribe Arabideae), the largest genus of the family with more than 400 species. In this genus the hybridization and the allopolyploidy have played an important rôle for the diversification of its species and their protection from cold stress, allowing the colonization of the high mountain alpine habitats and of Arctic. The instances of duplication of the genome have occurred in different geological age and have favoured the radiation, the ecogeographic diversification and, with the adaptation to new ecological niches, the increase of the species in most part of the genera of the tribes (Alysseae, Arabideae, Brassiceae, Cardamineae, Erysimeae, Heliophileae and Lepidieae).

And there are species living in aquatic environments, like the Subularia aquatica present with several subspecies in the Palearctic and in the Nearctic © Len Worthington
The radiation of the species of Lepidium in Australia took place in the Pleistocene (about 2 million years ago) due to climatic fluctuations that have caused the lowering of the temperature and the increase of the aridity creating new habitats and giving new ecological room to this genus that, thanks to the allopolyploidy, has been able to expand.
Usually, the polyploidization is followed by the diploidization (that include the genetic, epigenetic and transcriptional changes) which means that the state of the genome gets again similar to the diploid one, that is, more functional.
In the Brassicaceae are present annual, biennial, or perennial herbaceous plants, shrubs [Zilla spinosa (L.) Prantl], rarely lianas or trees. Usually, they are terrestrial plants, rarely aquatic (i.e. Cardamine lyrata Bunge). The rhizomes may be thin or thick and the stems simple or ramified, woody at times. In some genera are present glandular papillae multicellular or with tubercles (i.e. Bunias, Chorispora and Parrya).

Aliaria petiolata with flowers and terminal leaves. Brassicaceae have rosette basal leaves or are absent. Usually the cauline are alternate, simple or pinnate composite, palmate or bipinnate. Amplexicaul or absent petioles. No stipules and the foliar lamina margin can be entire, toothed, crenate, wavy, sinuate or doubly dissected © Dr. Giuseppe Mazza
The hairs (trichomes) can be unicellular, simple, pedunculate or star-shaped sessile, peltate or scaly.
The stems are usually erect, ramified or not, ascending, descending or prostrate, absent in Idahoa scapigera (Hook.) A.Nels. & J.F.Macbr.
The leaves distinguish in basal, in rosette, or absent, and cauline, usually present, normally alternate (opposite or spiralled in Cardamine angustata O.E. Schulz, C. concatenata (Michx.) O. Schwarz and in C. diphylla (Michx.) A.W. Wood and in Lunaria annua L.; subopposite in C. dissecta (Leavenw.) Al-Shehbaz and C. maxima (Nutt.) A.W. Wood and in Draba ogilviensis Hultén.), usually simple or compound, pinnate, palmate, or bipinnate.
The petioles can be amplexicaul, or absent, on whose base are at times present tiny glands. There are no stipules. The margins of the foliar lamina are variable, entire, toothed, crenate, wavy, sinuate or doubly dissected.
The inflorescences are terminal, usually racemose (often corymbs or panicles) or reduced to solitary flowers on peduncles forming at the axil of teh rosette-shaped leaves. The bracts are absent, present only rarely.
The Brassicaceae because of their homogeneity have, almost all, the same floral formula K 2+2, C 4, A 2+4, G 2 (superior).
The flowers have perianth with distinct calyx and corolla. They are bisexual, usually actinomorphic (weakly zygomorphic in Iberis, Ionopsidium, Pennellia, Streptanthus, Teesdalea); hypogynous, with 4 free sepals, in two crossed couples (decussate) on two cycles, usually deciduous, and merged at the base, saccate, tubular, campanulate. The corolla has 4 petals alternating with sepals, usually cross-shaped (cruciform), rarely rudimentary or absent. The stamens are 6, sometimes 2 or 4, on two verticils, usually 4 longer and 2 shorter, rarely of the same length. The staminal filaments are thin, merged at times, in pairs, rarely connate. The anthers have two thecae and open by means of longitudinal fissures freeing pollinic granules usually with three openings (3-grooved or tricolpate pollen) and with three nuclei (trinucleate).
At the base of the stamens can be present nectar glands variable in number, shape, dimensions and disposition, that secrete a sugary liquid collected in the saccate sepals.
The gynoecium is mi- or bi-carpellate with 2-locular superior ovary with false septum (replum), rarely 1-2ocular and without septum (Isatis, Clypeola), usually parietal placentation, rarely apical.

Heliophila coronopifolia. The flowers of the family are bisexual, usually regular. Corolla with 4 petals, alternate to sepals. The stamens are 6, at times 2 or 4, on two verticils, usually 4 long and 2 short, rarely equal © G. Mazza
The false septum originates at the margins of the carpels like outgrowths that merge and form the wall dividing the ovary in two cells following a longitudinal plane.
The ovary has 1 to 300 ovules, these can be anatropous or campylotropous and usually with very developed pluricellular nucleus (crassinucellate ovules).
The stigma is capitate or conical, entire or bilobate.
The pollination is mainly entomogamous, done by bees, flies, moths, coleopterans; in some species, if the pollinators’ visit is not sufficient, we have the self-pollination realized by the most external and shortest stamens. In some species the self-pollination is an obligation because the flowers do not open (cleistogamic flowers), like, for instance, Cardamine chenopodifolia Pers. that produces underground cleistogamic flowers; in others, but rarely, we have anemophily like Pringlea antiscorbutica R. Br. ex Hook. f.
The fruit is usually a capsule opening through 4 fissures placed two by two on the sides of the placentas (paraplacentar dehiscence). The two valves leave uncovered the replum on which are glued the seeds and this persists even after the dehiscence.
In Rorippa barbareifolia (DC.) Kitag. the valves are 3 to 6, usually 4, and in Tropidocarpum capparideum Greene they are 2 or 4.
The fruit of the Brassicaceae gets the name of silique if the length is thrice the width (Brassica, Sinapis, etc.), silicle if the length is less than three times (Lunaria, Lobularia, Biscutella, etc.). The diminutive silicle refers to the ratio between the two diametres and not to the size; in fact, many silicles stand among the largest of the family (Lunaria).
The fruits can have round section, (Brassica), flat, (Cardamine), or quadrangular, (Barbarea). They may also have the shape of samara (Isatis) or schizocarpic and, if they snap open, they throw the seeds far away (Cardamine). They can lose secondary the dehiscence and present narrowings that when ripe determine their fragmentation in articles with the seeds. This type of fruit takes the name of siliqua lomentacea (Raphanus). The silique can reduce to indehiscent fruit with two cells, lower and upper, fertile (Cakile) or, if the lower one aborts, the fruit becomes an achene (Crambe).
The fruit, as we can see, presents various morphological modifications thus it becomes an essential element for the determination at level of species, as well as the structure of the seed and of the nectariferous glands of the flower, even if they do not have an absolute value due to several exceptions. The seeds are small or of medium size, flat or roundish, winged or not, with narrow margins, ovoidal, oblong or globose, mucilagenous or less when wet.

Raphanus raphanistrum ssp. landra fruits with an enlarged detail. In the Brassicaceae they are usually dehiscent and when their length is at least three times the width they are called silicles. If then they have narrowings, like in this case, breaking when ripe, the fruits are scientifically called dehiscent lomentaceous siliques © Dr. Giuseppe Mazza
The seeds of Brassicaceae have two teguments, the outer one is multilayered, formed by an epidermis that varies in shape in the different species, with a subcutaneous layer of collenchymatic type and a deeper layer formed by cells having a thickened wall. The most internal tegument is usually thin and has a yellow or brown pigmentation that is maintained in the seeds. The embryo is usually curved, rarely straight. The cotyledons are entire, folded lengthwise around the radicle, spirally wrapped, or may have other arrangement. Usually, the seeds do not have endosperm, but, if present, it is monostratified and contains reserves (granules of aleurone and lipids). The germination is epigeal.
The embryo has two fleshy, at times oleaginous, cotyledons. For the systematics of many genera is important also the positioning of the cotyledons, that actually is the position of the radicle in respect to the cotyledon and this can be seen though the transversal section of the seed. If the cotyledons are flat, the embryo can have two positions: with the dorsal radicle stretched on the back of a cootyledon (incumbent cotyledon) and the embryo is called notorrizic or with radicle located against the margin of both cotyledons (accumbent cotyledon) and the embryo is called pleurorizic. If the cotyledons are folded to form a pipe where the embryo stands, we have the folded incumbent cotyledons and if, conversely, the cotyledons are folded on themselves, we have the conduplicate cotyledons.
Morphology and anatomy of the seed have an important taxonomical meaning in the Brassicaceae. Besides the reproduction by seeds (gamic or sexual reproduction), some species can reproduce also vegetatively. For instance, Cardamine bulbifera (L.) Crantz produces at the axil of the leaves, some globose bulbils, pyriform, violaceous that turn black and then fall down originating new plants in the following season.
The Brassicaceae systematics is continuously evolving, but during the last years have been published many molecular phylogenetic data and these results have solved many conflicts between the molecular data and the morphological ones.
Thanks to the molecular studies the family is divided up in two big groups, of which one is Aethionema (65 species), and the other represents the central nucleus of the family itself. This latter, in turn, is divided in three lineages. The lineage I includes 16 tribes, the lineage II has 6 tribes to which recently has been added the expanded lineage II with 21 tribes and the lineage III with 7 tribes.

When, like in Lunaria annua, the fruit’s length is less than three times the width, regardless of the size, they are called silicles. Here we note in transparency the seeds that some Brassicaceae can throw far away. Usually the fruits are capsules opening when ripe showing them sometimes stuck to their valves © Dr. Giuseppe Mazza
These lineages include the overwhelming majority of the species ascribed to the Brassicaceae.
To the lineage I belong the tribes: Alyssopsideae, Boechereae, Camelineae, Cardamineae, Crucihimalayeae, Descurainieae, Erysimeae, Halimolobeae, Lepidieae, Malcolmieae, Microlepidieae, Oreophytoneae, Physarieae, Smelowskieae, Turritideae, Yinshanieae.
To the lineage II belong the tribes: Brassiceae, Eutremeae, Isatideae, Sisymbrieae, Thelypodieae, Thlaspideae.
To the expanded lineage II belong the tribes: Alysseae, Anastaticeae, Aphragmeae, Arabideae, Asteae, Biscutelleae, Calepineae, Cochlearieae, Coluteocarpeae, Conringieae, Cremolobeae, Eudemeae, Heliophileae, Hillielleae, Iberideae, Kernereae, Megacarpaeeae, Notothlaspideae, Schizopetaleae, Scoliaxoneae, Stevenieae.
To the lineage III, the most evolved of the Brassicaceae, belong the tribes: Anchonieae, Buniadeae, Chorisporeae, Dontostemoneae, Euclidieae, Hesperideae, Shehbazieae.

Cakile maritima is a species of the Atlantic European dunes and the Mediterranean. The fruit is a conical-ovoid indehiscent silique, called biloment that separates when ripe in two articles containing one seed each © Harry Rose
The phylogenetic relations between these tribes are mainly based on sequences of nuclear ribosomial DNA ITS (Internal transcribed spacer) .
Here we describe the most taxonomically significant tribes.
I – Basal Group
Tribe Aethionemeae
This tribe is basal in rapport with all other Brassicaceae and is formed by only one genus Aethionema with 65 species mainly distributed in the Middle East and in East Europe. The centre of dispersion is in Turkey but with species extending eastwards, in Turkmenistan, and westwards, in Italy, Spain and Morocco.
Its species are found in the sub-Mediterranean garrigues and in arid meadows, on calcareous soild rich in skeleton (larger than 2 mm), very arid in summer, from the sea level up to the mountain belt. As an example, we cite Aethionema saxatile (L.) W.T. Aiton, perennial herbaceous plant of stony and rocky places of the hilly and mountainous plain of central-southern Europe, in North Africa and of western Anatolia.
To this tribe belong perennial herbaceous plants, often shrubs, occasionally annual herbs. Generally, the stems are numerous, they ramify from the base, forming a dense turf on dense bushes, often thorny, [Aethionema spinosum (Boiss.) Prantl]. The leaves are small, spirally arranged, entire, fleshy, sessile. The flowers, grouped in dense apical racemes, are numerous actinomorphic, of colour varying from white to clear pink or purple red. The fruit is a silicle, laterally compressed, in section of generally circular (suborbicular) shape, at times indehiscent and with wide wings and with entire or denticulate margin. Several species of the genus have seeds that, once wet, secrete mucilages to facilitate the adhesion to the ground. The presence of mucilages is also very important for the dispersion of the seeds and also for the colonization of new habitats.
II – Basal Group
Lineage I
Tribe Alyssopsideae
This tribe groups 9 specie, belonging to 4 genera (Alyssopsis, Calymmatium, Dielsiocharis, Olimarabidopsis), distributed in Afghanistan, Azerbaijan, Iran, Tajikistan, Turkmenistan and China.

Besides the seeds reproduction, some species of the Brassicaceae family can reproduce also vegetatively. Cardamine bulbifera, for instance, produces, at the axil leaves, bulbils globose, pyriform, violaceous , black when ripe, that later fall down originating new plants in the following season © Robert Hansmann and © Len Worthington
Olimarabidopsis cabulica (J. D. Hooker & Thomson) Al-Shehbaz et al., is an annual plant, about 12 cm tall present on the rocky slopes,in the valleys and banks of rivers at about 3000-4200 m of altitude. It has decumbent or erect stems, little ramified, pubescent with stellate trichomes. The basal leaves are subsessile, pubescent with oblanceolate, obovate or lanceolate foliar lamina, with entire margin, rarely toothed and acute apex. The cauline leaves are oblong or ovate, sessile, with auriculate base and entire or curved margin and acute apex. The sepals are oblong and pubescent. The petals, spatulate or oblanceolate, are attenuated at the base and have no claw, are of yellow colour fading when dried. The fruits are curved or straight siliques. The seeds are oblong, uniserial, of brown colour, in number of 5-18(-20) for each fruit. Blooming is vernal and the fruits are ripe between May and June. The base chromosome number is: x=24.
Tribe Boechereae
This tribe is almost exclusively of North America, and only one species, Boechera falcata (Turcz.) Al-Shehbaz, is found in the Russian Far East. It includes 9 genera and about 129 species, most of them belonging to the genus Boechera.

With unusual magenta red corollas, Boechera arcuata is native to California. Belongs to the tribe Boeachereae, with about 129 species mainly belonging to this genus © Mark Egger
All members of the tribe have mainly entire leaves (but Polyctenium and Sandbergia) and hairs (trichomes) ramified (absent in few species of Boechera, simple in the genus Nevada). Usually, they are perennial plants with well defined basal rosette. The cauline leaves, absent at times, are petiolate, sub-sessile or sessile. The base of the leaf is, or is not, auriculate, with usually entire or toothed margins. The racemes have no bracts with actinomorphic flowers with erect, ascending or expanded sepals, and the lateral couple, normally, is not saccate at the base. The petals are white, yellow, pink or violet. The pollen is 3-colpate ans the fruits are siliques, rarely silicles. The cotyledons can be either accumbent or incumbent. The base chromosome number is: x=7.
Tribe Cardamineae
This tribe of 15 genera and 417 species includes the genera Cardamine (239 species) and Rorippa (87 species). Both grow in all continents but Antarctica. The members of this tribe are mostly found in cool environments or aquatic and have leaves pinnately divided or compound (rarely palmate or simple).

Cardamine amara, common along water streams of Europe and west Asia from 600 to 2500 m of altitude, belongs to the tribe Cardamineae rich of 417 species © G. Mazza
They are annual, biennial, or perennial plants with hairs (trichomes) simple or absent and with racemose inflorescences usually with no bracts. The flowers are regular with erect, ascending, sepals and with the lateral couple only rarely saccate at the base. The petals are spatulate and of white, yellow, pink, lilac or violet colour. The pollen is 3-colpate. The fruits are siliques or silicles and can have a linear, ovoidal, ellipsoidal, pyriform form with smooth surface or with narrowings (torose). Usually, the cotyledons are accumbent (Cardamine amara L.), at times incumbent.
Cardamine acris Griseb. is a species endemic to Greece and is found in the humid meadows, in the rocky sites, in clefts, at the margin of woods at 1100-2100 m of altitude. It is a perennial, rhizomatous herbaceous plant, with erect stem, simple or ramified. The leaves form a basal hairy rosette, later on glabrous on the lamina but hairy on the margin. The leaves of the stem, mostly in the upper half of the stem, are pinnate or pinnatisect, similar to the rosette-like leaves, hairy on the margin. The sepals are ovate with membranous margins and the petals, obovate with a short claw, are of white, clear pink, to violet reddish colour. The stamens are 6, tetradynamous, with yellow anthers and stigma wider than the style. Blooming takes place from May to mid-July.

Cardamine pratensis shares the same range at lower altitudes. This genus is the most numerous of the tribe with more than half the species © Richard Collier
Tribe Halimolobeae
This includes 5 genera and 39 species exclusive to the New World. Most of them are located in central and norhern Mexico, three species in south-western USA and others in South America. The genera Mancoa and Pennellia have separate centres of distribution going from USA to Central America, from Bolivia to South America. The Halimolobeae are perennial or annual herbs with simple or ramified hairs, entire, sessile, leaves with auriculate base and entire, or rarely toothed, margin. The flowers, white, seldom violaceous, are grouped in racemes equipped with bracts; only two species of Mancoa do not have them. The seeds, if wet, are mucilaginous. The cotyledons are incumbent. The base chromosome number is x=8.
Tribe Lepidieae
This tribe is composed of 3 genera and 293 species. The main genus is Lepidium distributed in all continents but Antarctica. They are annual, biennial or perennial plants, rarely lianas or shrubs. They have simple trichomes or these can be absent. The cauline leaves are petiolate or sessile, with auriculate, or not, base, and margins usually entire or toothed.

Endemic to Spain Sierra Nevada, Lepidium hirum subsp. stylatum belongs to the Tribe Lepidieae. Grows in prairies, stony sites and humid zones from 2200 to 3400 m © Lucas Gutiérrez Jiménez
The racemes have no bracts, with actinomorphic flowers, sepals ascending up to spread apart, lateral couple with not saccate base. The petals are white, yellow, pink or violet. The fruits are generally dehiscent selicles, not segmented. The seeds are obovate-oblong, one per loculus, often mucilaginous. The cotyledons are flat, therefore they can be accumbent or incumbent. The base chromosome number is x=8.
Lepidium hirtum subsp. stylatum (Lag. & Rodr.) Thell. is a subspecies endemic to Sierra Nevada (Spain), that lives in the prairies, in the stony sites and in the humid zones over the 2.200 metres of altitude, up to 3.400 m. It is a perennial plant, up to 20 cm tall, with multistemmed woody rhizome, usually grey, pubescent. The stems are flexible, up to 20 cm tall. The leaves are basal, on long petioles, undivided, spatulate; the cauline ones have an ovate form of ca. 5-42 mm. The petals are white of ca. 3 mm. The stamens are 6 with violaceous-blackish anthers. The fruits are 4(5) mm silicles, hairy, with seeds of about 2 mm, ovoidal, smooth, of dark colour. This endemism is included in the red List of the Andalusian Vescular Flora.
Lepidium hirtum subsp. oxyotum (DC.) Thell. is a subspecies present in Corsica as well as in Crete in the uncultivated and rocky places in the mountains of the two islands. It is a pubescent-whitish plant, perennial, with short (8-20 cm) stems, reclinate and entire or toothed leaves.

Lepidium flavum. The genus Lepidium, present in all continents, but Antarctica, with its 259 species is the richest of the tribe Lepidieae © Mark Egger
The flowers, arranged on short racemose inflorescences, appear between May and July and present the calyx with hairy sepals and white petals. The fruits are silicles, oval-rounded, with wings entire or just jagged, and obovate seeds.
Lepidium flavum Torr. is an annual species native to California, Nevada and Mexico. It is met in the sandy alluvial desertic plains, along the brooks, from 600 to 1600 m of altitude. The stems are ramified and are generally prostrate. The cauline leaves have ovate to obovate to spatulate lamina; the basal leaves, arranged in rosette, have obovate to spatulate or oblanceolate lamina and entire margins. The flowers are grouped in corymbiform racemes slightly elongated in the fruit, with obovate-oblong sepals and yellow spatulate petals that, from March to June, colour the vast stretches of land where they grow. The stamens are 6 and the fruits produce ovate, 1-1,6 x 0,6-0,9 mm seeds having a peppery flavour.
Tribe Microlepidieae
This tribe includes herbaceous and shrubby plants distributed in 16 genera and 56 species typical to Australia and New Zealand. A distinctive characteristic of the tribe is given by some of its genera, such as Arabidella, Ballantinia, Pachycladon, and Stenopetalum, that, on the base of specific studies, descend from ancestors who have had duplication of the entire genome.

Murbeckiella boryi grows in cracks of siliceous rocks of Iberian Peninsula and North Africa at 1300-3300 m of altitude. The tribe Oreophytoneae, to which it belongs, hosts small perennial herbaceous plants with deep taproot roots. It has only 2 genera: Murbeckiella with 5 species and Oreophyton with just 1 species © Lucas Gutiérrez Jiménez
The genus Microlepidium, that gives the name to the tribe, regroups two species, M. alatum (J.M. Black) E.A. Shaw and M. pilosulum F. Muell. They are up to 20 cm tall plants, mainly erect, with simple and stellate hairs and often tangled branches. The leaves are obovate. Blooming occurs on spring with a long corymbiform inflorescence. The sepals are very small (about 1 mm) and the petals equal to the sepals, serrate or trilobate, of white colour. The fruit varies from cuneate to obcordate, 3-5 mm long, with serrate apex and with wings. M. pilosulum differs from M. alatum being hairy with winged fruits only in the third distal.
The genus Arabidella is formed by 7 species, typical of the semiarid regions of Australia. They are annual grasses or perennial shrubs, with erect posture, glabrous, papillose or with simple hairs. The leaves, petiolate, are entire up to 2 or 3 times divided in narrow segments. The inflorescences are dense or long racemes. The petals, with claws, are of a colour varying from yellow to white. The fruit is a linear silique, straight or slightly curved, dehiscent.
Tribe Oreophytoneae
The tribe includes the genera Murbeckiella and Oreophyton.

Physaria bellii is endemic to Colorado, where it grows from 1500 to 1800 m, in schistose and clayey soils. Belongs to tribe Physarieae that groups 7 genera and 183 species © Jeff Mitton
They are perennial herbaceous plants with deep tapering root, with simple, stellate, or very reduced, hairs. The stems are up to 25 cm tall or absent, of green colour, intense at times. The basal leaves are petiolate, rosette arranged, entire or subentire. The cauline leaves are pinnatifid or pinnatisect. The inflorescences are dense racemes with rather small and numerous flowers. The sepals are erect, the median ones obtuse, being the lateral acute. The petals, spatulate with claws, are white with violaceous shades, rarely pink. The stamens are tetradynamous. The fruits are generally curved siliques. The seeds, usually provided of wing of variable length, are brown or blackish.
Murbeckiella has 5 species native to the mountain zones of southern Europe, Caucasus and of north-western Africa. Murbeckiella boryi (Boiss.) Rothm. is a herbaceous plant with small white flowers and simple and dtellate hairs and is found in the shady sites, in the craks of siliceous rocks of the Iberian Peninsula and in North Africa at 1300-3300 metres of altitude. Murbeckiella zanonii (Ball) Rothm., endemic to the Romagnol Tuscan-Emilian Apennines, is a perennial herbaceous plant, 15-30 cm tall, with rather woody root and ascending-erect stems, pubescent, ramified at the top. The racemose inflorescences have white pedicellate flowers. It is found on rocks and siliceous substrata, on slopes and on escarpments, on screes and on loose stony debris, from 800 to 1500 m of altitude.

Cosmopolitan, at times infesting but with medicinal virtues, Capsella bursa-pastoris belongs to the tribe Camelineae © Dr. Giuseppe Mazza
The genus Oreophyton includes only one species, the O. falcatum (E. Fourn.) O.E. Schulz, native to Ethiopia and to tropical East Africa, where it grows near brooks, rocky ravines and on wet mud at 3900-4900 metres of altitude.
The base chromosome number is x=8.
Tribe Physarieae
The tribe is formed by 7 genera and 183 species.
The peculiarity of this tribe, compared to the others of the family, stands in the pollen that may have three, four or more openings (3-4-11-colpate pollen), whilst the remaining Brassicaceae have only three openings (tri-colpate pollen).
The tribe is present mainly in North America, and only some species of Physaria are separate in South America and one, P. arctica (Wormsk. ex Hornem.) O’Kane & Al-Shehbaz, is circumpolar.
They are annual plants, biennial or perennial, or small shrubs.
The cauline leaves are petiolate, sessile or subsessile, and the base of the lamina normally is not auriculate but the genus Paysonia. The margins can be entire, toothed or sinuate. Usually, the hairs are stellate and sessile whilst in Paysonias they are simple, forked and pedunculate.
The flowers are regular with erect sepals, enlarged, ascending or reflexed, with the lateral couple rarely saccate at the base. The petals are white, yellow, violet, orange.
The ovules are two or more per cavity and the fruits can be siliques or silicles, even swollen. The cotyledons are accumbent or incumbent.
The genus Physaria is the richest of species of the tribe (108 accepted species).
Physaria bellii G.A. Mulligan, species endemic to Colorado (USA), grows at altitudes between 1500 and 1800 metres on the schistose and calcareous soils of the geological formations Fountain, Niobrara and Pierre. It is a showy plant due to its flowers having sepals of pale yellow or yellow-green colour, and not clawed petals, spatulate or obovate (9-13 mm) of intense yellow colour.
The genus P. fendleri (A.Gray) O’Kane & Al-Shehbaz is of particular economic interest because of its seeds rich of oil, where the main constituent is the lesquerolic acid that is an important raw material for the production of cosmetics, resins, waxes, nylon, plastic, lubricating greases.
Most of the species of the tribe have base chromosome number x=8, but an aneuploid reduction has been reported at x=4 and an increase to x=11.

Smelowskia americana is an orophyte growing from 1600 and 4000 m of altitude. The Tribe Smelowskieae counts only 25 species assigned to this genus © Matt Lavin
Tribe Smelowskieae
This tribe includes 25 species of the genus Smelowskia, diffused in the boreal North America, Europe, central and eastern Asia. They are perennial plants, rarely annual, living in stony soils of moraines, on the cliffs or along the slopes. They have pedunculate hairs, usually stellate or bifurcate, at times also simple.The longest leaves are arranged along the base of the plant. The cauline ones, normally present, are petiolate, subsessile or sessile with not auriculate lamina at the base and usually lobate margins, at times entire or toothed. The inflorescence is a dense and spherical raceme that lengthens with the development of the fruits.The flowers are regular with upper sepals spread out and lateral couple not saccate at the base. The petals are white, pink, violet, at times yellow. The pollen is 3-colpate. The fruits are siliques or not segmented silicles, with four corners, containing many mucilaginous seeds. The cotyledons are accumbent or incumbent. The base chromosome number is x=6; polyploid x=12: aneuploid with x=11 and 10.
Smelowskia calycina (Stephan ex Willd.) C.A. Mey. is present in Asia and is found in the rocky slopes, in the gravel hills, crevasses and in the alpine meadows, from 2500 to 4900 m of altitude. They are 10-40 cm tall grasses, with simple stems, covered by simple or dendritic hairs, at times glabrous.

The tribe Erysimeae includes only the genus Erysimum with 274 accepted species. Erysimum rhaeticum is a plant endemic to the alpine arc found up to 1800 m of altitude and known as Swiss wallflower © Dr. Giuseppe Mazza
The cauline leaves are sessile, at times combed. The inflorescence is simple and bears flowers, between June and August, with persistent sepals and white or pale-yellow petals. The fruits are siliquae, usually ellipsoidal, that release, between July and September, seeds of brown colour.
Smelowskia americana (Regel & Herder) Rydb. Is found in America, a morphologically similar species, but with precociously falling sepals.
The Camelineae
This tribe has 8 genera and 36 species, mainly distributed in Asia and in Europe, with few species in North America.They are annual, without gland biennial plants. The hairs are pedunculate or sessile, dendritic or biforked, simple at times.
The cauline leaves are usually sessile, rarely with petiole and with usually entire margins. The base of the lamina is generally auriculate. The racemes have no bracts, with actinomorphic flowers. The sepals are erect, enlarged or ascending, with the lateral couple usually not saccate at the base. The petals are white, yellow, pink, lavender or violet. The pollen is 3-colpate. The fruits are dehiscent or indehiscent, silicles in Capsella, Neslia and Camelina, not segmented siliques, with circular or quadrangular section, in all other genera. The seeds are biserial or uniserial. The cotyledons are accumbent or incumbent. The base chromosome number is mainly x=8, reduced to x=5 in Arabidopsis thaliana, and x=4 in Stenopetalum.
Tribe Erysimeae
This tribe includes only the genus Erysimum with 274 accepted species. They are annual grasses, biennial or perennial, sometimes woody at the base and shrubby, with sessile hairs, usually stellate. The stem normally is very leafy, tetragonal or almost cylindrical. The leaves are simple, entire, pinnate with petiole or sessile. The inflorescences are racemose corymbs with numerous flowers. They are generally medium in size, and are of yellow, orange, rarely violet or white colour, with often short petioles. The sepals are erect, sacciform at the base. The petals, with claw, are usually longer than the sepals. The fruit is a silique with many seeds. The cotyledons are accumbent or, rarely, incumbent. The base chromosome number can vary from x=6 to x=11.
Erysimum cheiri (L.) Crantz is a species native to eastern Mediterranean, present in Italy as a naturalized exotic. It is a perennial plant with ramified stems, woody at the base, pubescent, with lanceolate, acute and entire leaves. The flowers, with 15-25 mm long petals and 8-11 mm sepals, are very fragrant and with colours varying from yellow to brick red. The fruits are siliques narrowed at the base, bicompressed with winged seeds.
Erysimum rhaeticum (Schleich. ex Hornen.) DC is a species endemic to the alpine arc found up to 1800 m of altitude, known as Swiss wallflower. It is a perennial herbaceous plant up to 70 cm tall, with erect stem and alternate leaves, entire, linear-lanceolate. The flowers, united in inflorescences, present yellow petals, 14-22 cm long and 7-10 mm sepals. Blooming occurs from April to June. The fruits are erect outspread siliques, slightly quadrangular, glabrescent at the corners. The seeds are 1,5-2,2 mm long.

The tribe Descurainieae has 6 genera and about 64 species, among which Descurainia bourgaeana here pictured at the Canaries © Jean Louis Verbruggen
Tribe Descurainieae
This tribe is composed by 6 genera and about 64 species distributed in North and South America, Eurasia, Africa, islands of Macaronesia.
The most representative genus is Descurainia. The tribe, that appears to be monophyletic, is formed by herbaceous plants or perennial (Descurainia is secondarily woody in the Canary Islands), with glands present or reduced (unicellular papillae). The hairs are pedunculate, dendritic or biforked, at times simple, rarely absent. Usually the cauline leaves are petiolate, with the base of the lamina not auriculate and usually toothed margins. The racemes can have bracts or may be without them, often elongated in the fruit. The flowers are regular, with erect sepals, ascending, enlarged or bent downwards (reflexed), with the lateral couple not saccate at the base. The petals are usually yellow, white at times, sometimes pink or violet. The pollen is 3-colpate and the fruits are seliques or silicles, dehiscent, not segmented, with roundish section. The ovary contains 4 to 100 or more ovules. Normally, the style is present and the stigma is entire. The seeds are usually biserial or uniserial, sometimes tetraserial (in Tropidocarpum). The cotyledons are incumbent, rarely accumbent. The base chromosome number is x=7. In Hornungia we have an aneuploid reduction with x=6.

The tribe Brassiceae has about 420 species, mostly Mediterranean. Cakile maritima, also present along the Atlantic European coasts, grows on the dunes © Sandy Steinman
Lineage II
Tribe Brassiceae
The tribe is composed of 53 genera and about 420 species that, excluding 5 species of Cakile of North America and of Central America, are typical of the Mediterranean area and the southern Asian one, though the range of the genus extends up to South Africa.
They are annual plants, biennial or perennial, without glands. The hairs (trichomes) are absent or are simple. The cauline leaves are petiolate or sessile, having the base of the lamina normally auriculate, with entire margins, toothed, jagged or pinnately lobate. Usually the racemes have no bracts, and are often elongated in the fruit. The flowers are regular with erect sepals, ascending or expanded. The two lateral sepals are saccate, or not, at the base. The petals are white, cream, yellow, pink, lilac, lavender or violet. The pollen is 3-colpate and the fruits are siliques or dehiscent or indehiscent silicles, often segmented, generally cylindrical (with quadrangular section in Erucastrum). The ovary contains 2 to 276 ovules and the stigma is entire or bilobate. The seeds are biserial or uniserial. The cotyledons usually are conduplicate, rarely accumbent or incumbent (Cakile).
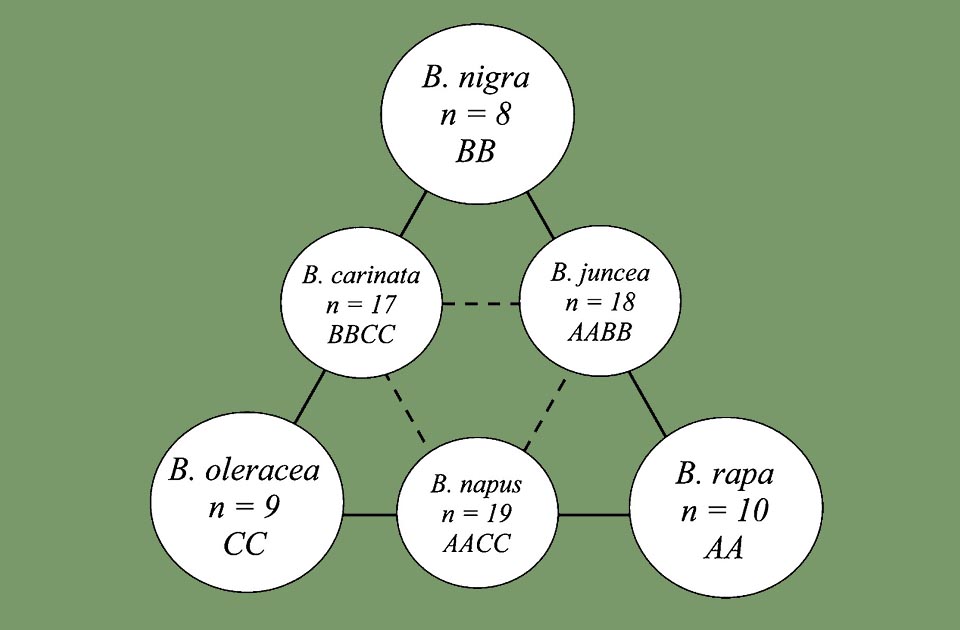
Nagaharu U. triangle diagram with genetic relations between 6 species of Brassica. In the corners the 3 progenitor species (B.rapa, B .nigra and B. oleracea) respectively with genomes AA, BB, CC, and on triangle sides the 3 allotetraploid species (B. juncea, B. napus and B. carinata). For each is shown the haploid chromosome number (n)
Cakile maritima Scop. is a succulent herbaceous plant diffused in the European Atlantic sandy coastal areas and in the Mediterranean Basin. The stems are ramified and prostrate with obovate to deeply pinnate alternate leaves. The fruit is a biloment, similar to a conical-ovoidal, but indehiscent, silique. When ripe it separates transversally in two articles containing one seed each.
The tribe includes various genera (i.e. Brassica, Raphanus, Sinapis) of huge importance in the human economy.
Woo Jang-choon (1898-1959), Korean-Japanese agronomist and botanist, accredited in the scientific literature with the name of “Nagaharu U”, in 1935 proposed a theory on the evolution and on the relations between the membres of the genus Brassica. The theory hypothesized that the genomes of three ancestral diploid species of Brassica had combined to create new tetraploid species and species of oily seeds cultivations. It is summarized in a triangle diagram showing the three ancestral genomes, named AA, BB and CC, placed at the corners of the triangle, and the three offshoots, indicated with AABB, AACC and BBCC, along its sides.

Very diffused in cultivation, Raphanus sativus var. sativus, tribe Brassiceae, exceeds 1 m with violaceous petals © Dr. Giuseppe Mazza
After this theory, the ancestors are:
– rapa L.(turnip) genome AB (n=10)
– nigra (L.) W.D.J. Koch (mustard) genome BB (n=8)
– oleracea L.(cabbage) genome CC (n=9)
Through natural crosses of these species, three new forms of allotetraploids were born:
– juncea (L.) Czern. (Indian mustard) genome AABB (n = 18) starting from B. nigra and B. rapa
– napus L. (rapeseed or oilseed rape) genome AACC (n = 19) from B. rapa and B. oleracea
– carinata A. Braun (Ethiopian rape) genome BBCC (n = 17) from B. nigra and B. oleracea
Tribe Eutremeae
The tribe includes the only genus Eutrema with about 42 species, all Asian and a few in North America. They are annual or perennial plants, without glands, with simple or absent leaves. The cauline flowers are petiolate or sessile with the not auriculate lamina at the base and entire or little wavy margins. The racemes have bracts, or do not have them, and are elongated in the fruit. The flowers actinomorphic with erect sepals and lateral couple not saccate at the base. Normally, the petals are white, rarely pink. The pollen is 3-colpate and the fruits are siliques or dilicles, dehiscent, not segmented, roundish pr quadrangular. The ovules vary from 2 to 96 per ovary. The seeds are uniserial, rarely biserial, and the cotyledons are usually incumbent. The base chromosome number is x=7.
Tribe Isatideae
This tribe is distributed mainly in central and western Asia, with a lesser number of species in Aurope. It counts more than 112 species regrouped in 6 genera. They are annual or biennial without glands, with simple or absent hairs. The cauline leaves are often sessile and with auriculate base of the lamina and with usually entire margins. The inflorescences are racemes, without bracts, and elongated in the fruits. The flowers are regular with erect or ascending sepals and lateral couples not saccate at the base. The petals are usually yellow. The pollen is 3-colpate and the fruits are siliques or silicles shaped like samarae, indehiscent, lignified. The ovules are 1 or 2 per ovary, the style is absent and the stigma is entire. The cotyledons are accumbent or incumbent.
The tribe gets its name from the genus Isatis, very known for its species I. tinctoria L., commonly known woad. From it, in the past, they extracted the blue dye, but in the XVI century with the advent of the indigo, a pigment gotten from Indigofera tinctoria L. and from I. suffruticosa Mill. with significantly hihgher yields, the woad was no longer used.
Tribe Sisymbrieae
This tribe is composed by only 3 genera and 52 species distributed in Europe and Asia, a few ones in Africa and one in North America. The tribe is monophyletic and sister group of the Schizopetaleae and perhaps also of the Brassiceae.

Not flowers, but a field of fruits: elegant flat silicles and trembling in the wind of Thysanocarpus radians, tribe Thelypodieae © Jeb Bjerke
They are annual or perennial plants without glands. The hairs are simple, ramified in Sisymbrium burchellii DC., or absent. The cauline leaves are petiolate or sessile with base of the lamina not auriculate and with margins generally lobate or toothed, entire at times. The inflorescences are racemose, usually without bracts, at times fasciculate, sometimes with solitary flowers (S. polyceratium L.). The flowers are regular with erect sepals and with lateral couple at times saccate at the base. The petals are yellow, sometimes pink or white. The pollen is 3-colpate. The fruits are dehiscent siliques, not segmented, usually with circular section. The ovary contains 6 to 120 ovules, sometimes even more. The base chromosome number is x=7.
Tribe Thelypodieae
The tripe regroups 29 genera and 287 species and is distributed in North and South America, with one species in the islands of the southern Indian Ocean (Pringlea antiscorbutica R.Br. ex Hook.f.). They are annual, biennial, perennial or shrubby plants. The hairs are usually simple, absent at times. The cauline leaves are petiolate or sessile with the base of the lamina auriculate or not, with entire margins, toothed or lobate. The racemes do not have bracts, are dense, with gyrophores often elongated in the fruits.

Yellow and white are the most frequent colours in the Brassicaceae family. Here Thlaspi arvense, species with garlic scent of the tribe Thlaspideae, with inflorescences rich of 30-70 candid flowers © Jean Louis Verbruggen
Usually, the fruits are regular, rarely zygomorphic, with erect sepals, enlarged or reflected, and with lateral couple, at times saccate at the base. The petals are white, yellow, orange, pink, lilac, lavender, violet, green, brown. The pollen is 3-colpate and generally the fruits are siliques, having circular section, or quadrangular, normally dehiscent. The ovary contains one to several ovules. The style is present or absent and the stigma is entire or bilobate. Usually, the seeds are biserial or uniserial.
The presence of P. antiscorbutica in the islands of the southern Indian Ocean (Kergeulen Islands, Heard and Mac Donald Islands, Crozet Islands, Prince Edward Islands) suggests that probably its ancestor reached America, about 5 million years ago, and settled in these islands, swept by very strong cold winds (150-200 km/h) thanks to the capacity of adopting the anemophily, and sometimes the self-pollination, due to the absence of pollinating insects. In fact, the flowers have very reduced petals and are grouped in large erect inflorescences that persist long time.
Thysanocarpus radians Benth. is a species native to central-northern California and to Oregon where it is found in the fields and in the humid meadows. It is an up to 60 cm tall annual grass with lobate leaves, about 5 cm long, and ephemeral bases and amplexicaule the caulines. The inflorescence is a raceme with whitish or violaceous small flowers. The fruit is a flat silicle,similar to a disc, hanging, glabrous or hairy. The wing covers the border of the disc and presents particular lines typical to the species.
Tribe Thlaspideae
The tribe is composed of 38 species regrouped in 12 genera mainly of south-western Asia, with a few ones in Europe. They are annual or biennial, rarely perennial, plants having no glands. The hairs are absent or are simple. The cauline leaves, petiolate or sessile, have a lamina normally auriculate at the base, and entire or toothed margins. The basal leaves often display palmate veins. The racemes usually do not have bracts, that elongate to the fruits. The flowers are regular with erect sepals and with lateral couple not saccate at the base. The petals are white, rarely pink or violet. The pollen is 3-colpate. The fruits are siliques or silicles, dehiscent, not segmented, with circular section or with four corners. The ovary usually contains 6 to 22 ovules, rarely many more. The stigma is entire. The seeds are uniserial, streaked or coarsily reticulate.
Thlaspi arvense L is an annual species; 30-60 cm tall, that smells of garlic. The stem is erect, angular, usually simple. The cauline leaves are lanceolate, lightly spatulate, 2×6 cm large with a usually toothed margin, sessile,with sagittate base. The basal leaves are petiolate,oblong with entire margin. The inflorescences, formed by 30-70 white flowers, are long racemes.
Alliaria petiolata (M. Bieb.) Cavara & Grande is a perennial plant, more than one metre tall, almost glabrous that, if rubbed, emits a very persistent smell of garlic. This smell persists also on the milk of the cows that have eaten it.

Odontarrhena alpestris. The tribe Alysseae includes 25 genera and 306 species mainly belonging to the genera Alyssum and Odontarrhena © Linda and John Reinecke
Expanded lineage II
Tribe Alysseae
The tribe is formed of 25 genera and 306 species, most of them falling in the genus Alyssum (116 accepted species) and Odontarrhena (88 accepted species).
They are annual, biennial or perennial plants, characterized by stellate hairs, pedunculate or sessile or biforked, at times mixed with the simple ones.The cauline leaves are petiolate or sessile with the base if the foliar lamina not auriculate and with the margins usually entire. The inflorescences are racemes generally without bracts. The flowers are actinomorphic with erect, open wide, sepals, whose lateral basal couple is rarely saccate. The petals are white or yellow, at times orange, pink or violet. The stamens are 6, four equal and two smaller (tetradynamous). The ovary is superior, bicarpellate, with 2-16 ovules and with usually entire stigma, rarely bilobate. The fruits are dehiscent, not segmented, siliques. The seeds are biserial, rarely uniserial. This tribe is met in Europe, in western Asia, and in north-western Africa.

Nebrodi Alyssum (Odontarrhena nebrodensis) is an endemic species present, over the 1200 m, on gravelly slopes and rocky and calcareous of the Madonia, the short mountain ridge at the centre in the northern part of Sicily (Italy) © Salvatore Cambria
The Alpine madwort Odontarrhena alpestris (L.) Ledeb., present in Italy, is a perennial plant of cespitose, woody, look, ramified from the base, with yellow flowers in dense inflorescences in apical racemes.
The Alyssum nebrodense (Odontarrhena nebrodensis (Tineo) L. Cecchi & Selvi) is a species endemic to the Madonie (Sicily), that grows on the gravelly and calcareous rocky slopes, over the 1200 m of altitude. It is a perennial plant, 3-16 cm tall, with woody, twisted and greyish stems, and with ascending or erect floriferous branches. The leaves, often glabrescent, are linear-spatulate, the basal being smaller than the cauline. The racemes are umbelliform with yellow (2-2,5 mm) flowers and that from during the months of May and June. The fruits are silicles.
Tribe Anastaticeae
The tribe counts 14 genera and about 80 species typical of Africa and of south-western Asia.
They are annual or perennial grasses, at times shrubs, rarely small trees, without glands. The hairs are sessile, stellate or exclusively unicellular with one biramous section. The leaves have short or sessile petioles, and generally entire lamina, not auriculate at the base. The inflorescences are racemes without bract, with gyrophore that elongates during the fruitification. The flowers are actinomorphic, quite rarely zygomorphic, with extended or erect ascending sepals and lateral couple usually not sacciform at the base. The petals are white, pink, violet, yellow or orange or brown.
The pollen is 3-colpate. The ovules vary from 2 to many. The fruit is a silique or a silicle, dehiscent, with circular or quadrangular section. The style is well differentiated and the stigma is entire or lobate. Unserial seeds, with or without wings.
In addition to the aforementioned Anastatica hierochuntica whose genus has given the name to the tribe, we mention Lobularia maritima (L.) Desv., called “common filigree”or “sweet alyssum”. It is a perennial plant, cespitose, 15-10 cm tall, frequent in the back dune zones but that can also go to locations far away from the sea, up to 1200 m of altitude. It has taproot roots srom which emerge stems, woody at the base, ascending and ramified. The leaves are tomentose, simple, of green-greyish colour. The basal ones are spatulate, whilst the cauline are alternate lanceolate-linear. The flowers, regrouped in racemose inflorescences, have calyx with 4 very small (1 mm) sepals, and corolla with 4 2-3 mm petals, generally of white or yellowish white colour, at times mottled pink. The stamens are tetradynamous with yellow anthers. The ovary is superior syncarpic bicarpellate and the stigma is capitate and persistent. The fruits are elliptical silicles of yellow or red colour. The seeds, small (1-1,5 mm), and one per cavity, are ovate, brown-reddish, without wings or, if these are present, very narrow. It is a reflowering plant sometimes utilized as ornamental plant.

Darba cretica is a small perennial herbaceous plant of the tribe Arabideae. Endemic to the Crete mountains, it grows on clefts and rocky ledges, stabilized stony and gravelly at 1100-2400 m of altitude. It is characterized by small hairy eaves in basal rosette with entire or toothed margin and flowers with yellow petals © Salvatore Cambria
Tribe Arabideae
This tribe includes 18 genera and more than 600 species, mainly distributed in Eurasia, in America and in Africa. The tribe is formed by the genus Draba, the richest of species (402 accepted), but with characteristics very different from each other. They are annual, biennial, or perennial plants, having no glands, with simple or biforked hairs, stellate, dendritic, cruciform, pedunculate or sessile. The cauline leaves may be petiolate or sessile with base of the lamina auriculate or not and with entire or toothed margins. The racemes, normally devoid of bracts, often elongate during the fruitification. The flowers are regular with erect sepals, enlarged, ans with the lateral couple rarely saccate at the base. The petals are white, yellow, orange, pink or violet. The pollen is 3-colpate and the fruits are siliques or silicles normally dehiscent, not segmented with circular section. The ovary contains 2 to 70, or more, ovules. The stigma is entire, rarely bilobate. The seeds are uniserial or biserial.
Draba cretica Boiss. & Heldr. is a tiny perennial herbaceous plant, endemic to the mountains of Crete, characterized by small simple leaves, hairy, in basal rosettes, ovate, with entire or toothed margin and flowers with yellow petals.

The tribe Biscutelleae has only 2 genera: Biscutella and Megadenia. Here Biscutella laevigata, a perennial herbaceous species frequent in Europe up to 2300 m of altitude, with its typical discoid silicles wind disperses © Dr. Giuseppe Mazza
The fruits are compressed silicles, elliptical or oblong with stellate hairs. This species lives in clefts and on rocky protrusions, on stony soils and on stabilized screes at an altitude at an altitude of 1100-2400 metres.
Tribe Biscutelleae
The tribe is formed by 2 genera and 60 genera, distributed in Europe, northern Africa, and in south-western and eastern Asia. The genus Biscutella L. with its 57 species, present in Europe and in the Mediterranean area, is the most represented, whilst the genus Megadenia Maxim. itself, has only 3 Asian species.
They are annual or perennial herbaceous plants, at times lightly suffrutescent, leaves basal in rosette and alternate cauline on the stem, simple to pinnatifid, with entire or toothed margins. The flowers, actinimorphic and bisexual, are grouped in terminal racemes. The sepals are free and petals of yellow colour are spatulate with claw. The ovary is superior. The fruits (silicles) are formed by two discoid spaces merged on a side, with protruding style and cilia on the margins.
Biscutella levigata L. is a perennial herbaceous species, upto 50 cm tall, that, in its various subspecies, is found in good part of Europe, on stony substrata, arid open meadows, at times also along the edges of mountain roads, from the sea level up to 2300 m of altitude. It is an up to 50 cm tall plant, with taproot rhizome and stems woody at the base and ramose at the top. The basal leaves are arranged in rosette, linear, spatulate with margin at times jagged. The cauline ones are smaller, sessile, of green colour, greyish due to the rich down. The inflorescence, formed by pedunculate flowers, is a low or compact raceme.The flowers appear fom April to August and display four yellowish sepals and four 5-10 mm yellow petals. The fruit is a glabrous silicle of 5,5 x 9-13 mm, formed by two discoid spaces. When ripe, the silicles are dispersed by the wind to then free small brown seeds, flat and reniform.
Tribe Cochlearieae
This tribe present in Europe, north-western Africa, northern Canada, Alaska and northern Asia, counts two genera: Cochlearia L. (16 species) and Ionopsidium Rchb. (8 species).
They are annual, biennial or perennial glandless plants. The hairs are absent. The cauline leaves are petiolate or sessile with the base of the lamina auriculate or not and with margins generally entire or toothed. The racemes are bractless that elongate during the fruitification. The flowers are regular, rarely zygomorphic, with ascending and open wide and lateral couples not saccate at the base.

Spectacular blooming of Cochlearia officinalis, tribe Cochlearieae, at Capo Lizard, in Cornwall © Philip Goddard
The petals are white and have a claw. The pollen is 3-colpate. The fruits are silicles, dehiscent, ovoidal. The ovary contains 8 to 14 (or many) ovules. The stigma is entire. The seeds are biserial with the cotyledons usually incumbent, that is, in the seed the radicle leans towards the cotyledon.
Ionopsidium savianum (Caruel) Ball ex Arcang. is present in central-northern Spain and in central Italy. It is an annual herbaceous plant, with erect posture, up to 10 cm tall. The hairs, basal, with 6-10 mm petioles, are spatulate with lanceolate lamina: the cauline ones are amplexicaul whose dimensions slightly reduce. The flower arranged in aphyllous racemes, have sepals averagely 1,5 mm long and white 3-4 mm petals, the outer ones slightly bigger than the others. The style is persistent, about 1 mm long. The fruits are winged silicles bigger than the others, often of violet colour and contain 2-3 seeds.
Tribe Coluteocarpeae
This tribe, present in Eurasia, North America and Patagonia, counts 13 genera and 151 species.

Noccaea rotundifolia is a perennial species endemic to alpine arc, with stoloniferous stems infiltrating screes and loose debris from 1600 to 3200 m of altitude © Olive Titus
The genus with more species is the Noccaea (92 species), followed by Thlaspiceras (11 species), Microthlaspi (5 species) and Pseudosempervivum (5 species). The remaining genera count from 4 to 1 only species.
They are annual, biennial or perennial plants with simple or stoloniferous stem, generally erect, more or less ramified. The basal leaves, arranged in rosette, are spatulate or lanceolate, at times coriaceous and rigid, often glossy, that become thinner on the petiole with entire, serrated or toothed. The lamina of the cauline leaves has auriculate base. The flowers, actinomorphic, are grouped in racemes, usually corymbs, that elongate or not during thr fruitification. The sepals are erect, oblong or ovate, with lateral couple not saccate at the base. The petals, white, pink or violet are spatulate sometimes linear, longer than the sepals, with well evident claw and lamina with obtuse or rounded apex. The stamens are lightly tetradynamous with filaments not enlarged at the base. The anthers are ovate with nectariferous glands. The fruits are compressed silicles, angustisette, dehiscent. The ovary is superior with 4-14 ovules. The stygma is capitate. The seeds are lightly compressed, winged or not, ovoid. The seeds are finely reticulate, at times smooth, not mucilagineous if wet. The cotyledons are accumbent. The base chromosome number is x=7.

Conringia austriaca, tribe Conringieae, amazes due to the large leaves wrapping the stem and the inflorescence © Hermann Falkne
Noccaea corymbosa (J. Gay) F.K. Mey. is found in the western part of the French, Italian and Swiss Alps, on stony humid soils, on rubble and moraines, generally of siliceous nature, at altitudes above the 3000 m. It is a perennial herbaceous plant with fleshy leaves forming thick and dense tufts. The inflorescence is a short raceme with very ornamental flowers, of dark pink-violet colour. The fruits are 8-12 mm compressed silicles, dehiscent, containing 4-9 0,8-1,2 mm long seeds.
Noccaea rotundifolia (L.) Moench is a specis endemic to the alpine arc. It is a perennial plant with stoloniferous stems that penetrate the screes and the rubbles, usually of siliceous nature, at an altitude of more than 3000 m. It is a perennial herbaceous plant with fleshy leaves forming thick and dense tufts. The inflorescence is a short raceme with very ornamental flowers, of pink-dark violet colour. The fruits are compressed 8-12 mm silicles, dehiscent and contanining 4-9 seeds long 0,8-1,1 mm.
Tribe Conringieae
This tribe is formed by 9 species, belonging to 2 genera (Conringia and Zuvanda), diffused in Europe and in the Middle East.

Also in Conringia orientalis, called Hare’s ears mustard, the flashy and waxy leaves wrap the stem. The petals at the base have thin claws © Ori Fragman-Sapir – www.flora.org.il
They are annual plants, at times biennial, without glands. The hairs are absent, or, if present, simple. The cauline leaves are sessile with base of the lamina cordate, auriculate or, rarely, sagittate, and with entire margins, rarely crenulate. The inflorescences, bractless, are racemose. The flowers are regular, with erect sepals and lateral couple saccate at the base. The petals are white or yellow, with claw. The pollen is 3-colpate and the fruits are dehiscent siliques, not segmented, generally with circular section.
The ovary contains 10-50, or more, ovules. Normally, the stigma is capitate and entire. The seeds are uniserial wih cotyledons usually incumbent. The base chromosome number is x=7 or, rarely, x=9.
Conringia austriaca (Jacq.) Sweet is a species present in Crimea, Caucasus and Asia Minor and in south-eastern Europe (Bulgaria, Romania, Greece, Appenninic Italy), it snorthern bounday, and in Austria. It is found in the xerothermic prairies and in the woody steppes, only on clayey dry sandy soils from the plains to the hills. It is an annual to biennial plant, up to 100 cm tall, with a scarcely ramified, at times, stem. The basal leaves, on short petioles, are ovate with entire margins, usually dry at the moment of the blooming. The cauline leaves, also with entire margins, are 6-8 cm long, ovate and all weakly wrapping the axis.

ative to eastern Mediterranean, Iran, Iraq and Arabian Peninsula, Zuvanda crenulata, tribe Conringieae, usually grows in sandy areas and abandoned fields © Ori Fragman-Sapir – www.flora.org.il
The inflorescence forms from April to May and is a cluster that elongates with the ripening of the fruit. The flowers have a 5-6 mm calyx and a corolla with 6-8 mm petals, of lemon yellow colour, being the external ones concave. The flowers are octagonal erect silicles that ripe 2,8-3 mm long seeds, of dark brown colour. It is a fairly rare and endangered species, protected in some Nations, already extinct in the Czech Republic.
Conringia orientalis (L.) C. Presl, called hares’ ear mustard, is native to Europe and to Asia, but introduced by man in North America, Mexico, north-western Africa and in Australia where, in the plains and in the prairies, is an invasive species. It is an annual grass that produces an erect stem, little ramified, 30-70 cm tall.
The leaves, wrapping the stem, are fleshy and waxy, the 5-9 cm basal ones, oblanceolate to obovate and the 3-10 cm sessile, lanceolate cauline ones. The inflorescences appear from April to August and bear flowers with pointed sepals and 7-12 mm petals, provided of thin claws, of lemon yellow or pale yellow colour. The fruit is an erect-patent silique, linear, up to 13 cm long. The 2-3 mm seeds are oblong of light brown colour. The plant is toxic to livestock.

Menonvillea cuneata is and Andean orophyte, tribe Cremolobeae, growing between 1700 and 4600 m of altitude © Claire De Schrevel
Zuvanda crenulata (DC.) Askerova is a species native to eastern Mediterranean, to Iran, Iraq and Arabian peninsula. It generally grows in sandy areas, in abandoned fields, from 300 to 1000 m of altitude. It is an annual herbaceous plant, ramified at the base, with an up to 25 cm tall stem. The leaves are linear and the flowers are present from December to April. The petals measure 16-23 mm and are of violet colour. The fruits are tuberculate and glabrous siliques.
Tribe Cremolobeae
In this tribe stand 3 genera and 36 species disributed in South America along the Andes from northern Argentina and from Chile up to northern Colombia.It is a tribe formed by three genera (Cremolobus, Aimara and Menonvillea), morphologically defined by the indehiscent schizocarpic silicles.They are grasses, annual or, more commonly, perennial, rarely lianas, glandless, with simple or ramified caudex from which emerge stems, one to many, usually simple, erect or decumbent. The hairs are normally simple. The basal leaves are absent or, if present, simple, entire or toothed, sessile or petiolate, arranged in rosette or not, pinnate or lobate palmate or combed.

Menonvillea nordenskjoeldii is a species endemic to Patagonia, Argentina and southern Chile growing on rocky soils between 500-2100 m of altitude © Rien Pieterman
The cauline leaves are missing or, if present, are petiolate or sessile, not auriculate at the base, entire, trifid, pinnatifid orcombed. The racemes are terminal, less freauently, axillar, like corymbs, with or without bracts, elongated or not in the fruits. Usually the sepals are ovate or, at times, oblong, free, deciduous, erect, with both couples saccate or not. The petals are yellow, yellowish-green, white or cream white, longer than the sepals with claw at times differientated fom the lamina. The stamens are 6, erect, subequal or tetradynamous. The anthers are ovate or oblong, generally with nectariferous glands. The ovary has two ovules with subapical placentation. The silicles are schizocarps that break when ripe in 2 oblong, ovoid, orbicular or globose mericarps, dorsally compressed, not segmented, with one seed each. The replum is persistent and thickened. The stigma is capitate and the cotyledons are incumbent or accumbent.
Menonvillea cuneata (Gillies & Hook.) Rollins is a species endemic to Andes, between Chile and central Argentina. It grows on stony and rocky mountain slopes, at altitudes between 1700 and 4600 m. It is a perennial herbaceous plant with ascending stems, straight, 10-20 cm long, provided of hairs, simple or ramified, at times woody at the base. The leaves, cuneate at the base, are sessile or poorly petiolate, oblanceolate, 10-20 mm long and 4-10 mm broad. The inflorescence is terminal and elongated in the fruit.

Xerodraba lycopodioides belongs to the tribe Eudemeae that includes about 36 species distributed along the Andes. Endemic to Patagonia, challenges the rigours of the cold steppe between 100-1500 m of altitude © Rien Pieterman
The sepals are 4-5 mm long, hairy and deciduous. The corolla has spatulate petals, 6-10 mm long, generally white, reddening when withering. The fruits are ellipsoidal siliques, winged, glabrous and hairy.
Menonvillea nordenskjoeldii (Dusén) Rollins is an endemic Chilean species present in the Region of the Lakes and in northern Patagonia, in a humid climate, on screes at altitudes between 900 and 1700 metres. It is a perennial plant with fleshy rhizomes, up to 60 cm long, forming 10-30 cm tall stems. The leaves are glabrous, hairy at times. The inflorescence is dense and spherical formed by white flowers appearing from late spring to summer.
Tribe Eudemeae
This includes about 36 species distributed along the South America Andes, from southern Colombia up to southern Chile and Argentina. In turn, the tribe is divided in two main different lineages due to their geographical distribution and their climatic niches: the Andean lineage, centre-northern, includes the genera Aschersoniodoxa, Brayopsis, Dactylocardamum and
Eudema, and the Southern Andean lineage of Paraguay that includes the genera Alshehbazia, Delpinophytum, Onuris and Xerodraba.
They are perennial herbaceous plants, glabrous or pubescent, without glands. The hairs, on short peduncles, are biforked or simple. The basal leaves are sessile, rosette-like, not auriculate, entire or toothed. The leaves of the stem are absent. The flowers are arranged in racemes, bractless, that elongate during the fruitification, or are solitary. Generally, they are regular with erect or open wide sepals and with the lateral couple not sacciform at the base. The petals are white or yellow and the ovary contains 2 to 60 ovules. The fruits are dehiscent, siliques or not segmented silicles. The style is generally very short and the stigma is entire. The cotyledons are incumbent.
Tribe Heliophileae
This tribe is formed by only one genus (Heliophila) with about 107 species endemic to South Africa, mostly confined in the south-western part. Most of them are met in two hotspots of biodiversity: the Cape Floristic Region, the smallest floral kingdom on the earth, with mostly winter rainfalls, and the Succulent Karoo, the semidesertic region of the Afrotropical realm. Fewer species can be found in the rest of southern Africa: Namibia, Swaziland and Lesotho.
The tribe counts annual herbaceous or perennial shrubby plants, at times even lianas, usually glabrous or with simple hairs. The leaves are variable, narrow or wide, but usually simple and can be entire, lobate or pinnatipartite.

Iberis odorata, tribe Iberideae, is native to the Canary Islands, North Africa, central Greece up to Iran © Ori Fragman-Sapir – www.flora.org.il
The inflorescences are racemose, and when ripe, they produce dehiscent fruts. This tribe has species standing among the most diversified in the family of the Brassicaceae. As a matter of fact, we note various colours of the flowers, such as blue, white or pink, the most common. Blue is an unusual colour and is found only in another genus, genetically distant, and peculiar to Himalaya: Solms-laubachia of the tribe Euclidieae. Also the size of the flowers varies remarkably. Species do exist, like H. juncea (P.J. Bergius) Druce, with quite big petals of pink, violet or white colours and species such as H. pectinata Burch. ex DC., with very small petals, just 1,2 mm long, of white colour.
Tribe Iberideae
This tribe is composed by two genera: Iberis with about 27 species and Teesdalia with 3 species. It is mainly distributed in Europe, with few species in north-western Africa, in Turkey and in south-western and central Asia.
They are annual or perennial plants, glandless. The hairs are simple or absent. The cauline leaves are petiolate or sessile, at times they can be absent. The base of the foliar lamina is not auriculate and the margins are generally entire, at times toothed or lobate. The inflorescences, bractless, are racemes that usually get longer during the fruitification. The flowers are zygomorphic, rarely actinomorphic with erect sepals, ascending or expanded, and with lateral couple not saccate at the base. The petals are white, pink or violet, with claw. The pollen is 3-colpate. The fruits are dehiscent silicles, oval and at times winged. The ovary contains 2 or 4 ovules. The style is habitually evident (sometimes absent in Teesdalia). The stigma is entire or with 2 lobes. The seeds have accumbent cotyledons.
Iberis procumbens Lange is a species endemic to the Iberian Peninsula. It grows mainly on coastal sands and dunes, at times even on cobblestones and stony soils. It si a perennial herbaceous plant with stem, reclined or erect, apically ramified. It has glabrous or hairy leaves, at times ciliate at the margin. The basal ones are spatulate or ovate, 10-20 mm long, fleshy, entire or with small teeth borne by petioles. The cauline leaves are oblanceolate or spatulate, 10-15 mm long. The inflorescence is corymb terminal with flowers having calyx with ovate sepals and corolla with 5-9 mm long petals, white or pink, the outer ones being larger. The fruits are merged silicles.
Two subspecies do exist: I. procumbens subsp. procumbens Lange the typical subspecies present on the Artlantic coasts of the Iberian Peninsula, particularly in those of the Galician and Portuguese ones, and I. procumbens subsp. microcarpa Franco & P. Silva, typical to the coastal or sub-coastal hills mainly calcareous, pebbly soils: 50-400 m of the central Portugal.
Iberis sempervirens L. is a species native to central-southern Europe, North Africa ans western Asia. It is a small, 30 cm tall shrub, present in lightly shaded sites, on sandy soils at times clayey, rich of nutrients. The leaves are alternate, simple, linear-oblong, coriaceous, of dark green colour. The flowers are white grouped in racemose inflorescences that cover all the plant and take form in spring up to early summer.

Iberis sempervirens, tribe Iberideae is a small 30 cm tall shrub of central-south Europe, North Africa and west Asia. The candid inflorescences grouped in racemose inflorescences cover all the plant in spring up to early summer © Dr. Giuseppe Mazza
The fruits are roundish oval silicles that free 2-3 mm winged seeds.
Tribe Schizopetaleae
In the past, this tribe included six genera, Caulanthus, Mathewsia, Schizopetalon, Sibaropsis, Streptanthella, Werdermannia but thanks to the recent molecular studies, supported also by the geographical distribution and by the morphology, the number has been reduced to two: Mathewsia (7 species), and Schizopetalon (10 species). These two genera are mainly distributed in the southern desert of Atacama and in the Matorral of central Chile. Some species can be found in southern Peru and in the neighbouring mountain areas of Argentina.
The genus Mathewsia includes perennial grasses or shrubs, with simple and dendritic hairs and with fragrant flowers having long white corolline tubes and reticulate pollen grains for the pollination by means of moths. Schizopetalon is morphologically distinguished from Mathewsia because of the annual posture and the presence of white pinnatifid petals having 7-9 linear to strictly oblong lobes that visually appear as unevenly cut. The name of the genus had been rightly given considering the peculiarity of the flower. The seeds are mostly uniserial and the cotyledons are filiform or incumbent.
Tribe Stevenieae
Tribe recently established formed by two genera: Macropodium (2 species) and Stevenia (8 species).
It is considered sister group of the tribe Arabideae. The whole tribe is present in China, Japan, Kazakhstan, Mongolia, Russia, whose membres live along the streams, the humid shady slopes, the rocky cliffs, from 100 to 1700 m of altitude. This tribe has a ring-shaped phytogeographic distribution around the system of high Asian mountains, interrupted and separated by genetically distinct membres belonging to the sister tribe Arabideae with the genera Baimashania, Pseudodraba and Sinoarabis.
They are herbaceous plants, annual, biennial or perennial, sometimes rhizomatose with simple, biforked or stellate hairs. The stems are erect, not ramified or ramified in the upper half. The basal leaves, normally arranged in rosette and petiolate, are simple, entire, jagged or toothed and the cauline leaves are entire, sessile or with petiole. The racemes are numerous with few flowers with or without bracts, they elongate during the fruitification. The sepals are oblong-linear with the base of the lateral couple not saccate. The petals are white, pink, violet or yellowship with ovate-shaped laminawith claw differentiated from the lamina, shorter than the sepals. The stamens are 6 tetradynamous with filaments dilated at the base and ovate or oblong-linear anthers. The nectariferous glands are 2 or 4 lateral, absent the median glands. The stigma is sessile, captate, entire. Every ovary contains 8-22 ovules (Macropodium) or (2-)4-40 in Stevenia. The fruits are siliques or dehiscent silicles with replum having complete septum, membranous, translucent. The seeds are uniserial, winged or not, ovate or oblong, often flattened, not mucilaginous if wet and with accumbent cotyledons. Base chromosome number x=15.

Called sea stock or wallflower of the dunes, Matthiola sinuata has a Mediterranean.Atlantic distribution typical of the sea dunes © Robert Combes
Lineage III
Tribe Anchonieae
This tribe includes 10 genera and 75species distributed in Europe, Asia and Africa.
They are annual, biennial or perennial plants, normally with multicellular glands and simple hairs, pedunculate, dendritic and biforked, sometimes mixed with the simple ones. The cauline leaves are petiolate or seddile with the base of the lamina not auriculate and with margins usually entire, toothed or pinnatisect. The racemes have no bracts and are often elongated in the fruit. The flowers are actinomorphic with erect sepals and wuth the lateral couple saccate at the base. The petals are white, yellow, pink, violet. The pollen is 3-colpate. The fruits are siliques, not segmented, dehiscent with circular or four-cornered section. The ovary contains 15 to 60 ovules. The style is well evident and the stigma has two lobes, often connivent. The seeds are uniserial, usually with accumbent cotyledons.
The base chromosome number is x=7, excepting the genus Matthiola that has chromosome variations from x=5 up to x=8.

Striking image of Matthiola arabica in the desert and close-up, below, of the odd rolled ripening silique © Ori Fragman-Sapir – www.flora.org.il
Matthiola arabica Boiss. is an Arab-Saharan species that is found in semidesertic or desertic areas in stony and sandy soils. It is a perennial plant, of greyish colour due to the presence of many hairs, with straight and very ramified stem. The flowers appear from March to May and have four long petals with wavy border of pale or dark pink colour. It is an allelopathic species, which means that is capable to emit toxic substances in order to hinder the germination and the growth of other species to defend its own ‘vital space’.
Matthiola maderensis Lowe, species endemic to Madeira, is an up to 1 m tall biennial or perennial herbaceous plant. It grows on cliffs, on coastal rocks and on dry soils. It has an erect stem, usually ramified at the base, with basal lanceolate leaves, entire, hairy, 5-25 cm long, of silvery green colour. The flowers, very fragrant and arranged in terminal racemes, are tetramerous, with erect sepals and obovate petals, 1,8-2,8 cm long, of violet, purple or, rarely, white colour. The fruit is an 8-15 cm long silique.
Tribe Buniadeae
This tribe counts only one genus with two species Bunias erucago L. and Bunias orientalis L. diffused in Europe and in Asia. They are biennial or perennial grasses, 40-100 cm tall, scarcely to densely hairy with simple and ramous, pedunculate glandular hairs.

Not less strange is the symmetrical silicle of Bunias erucago, tribe Buniadeae © Robert Combes
The stems are erect, ramous at the top, rough and tuberculate. The seeds are subglobose, brown-reddish with conduplicate cotyledons. The upper branches of the inflorescence are reddish and covered by glandular multicellular hairs. The basal leaves are arranged in rosette, spatulate with 5-7 little accentuated teeth on each side (B. orientalis).
The cauline leaves are oblong-spatulate with more or less toothed margins. The sepals are saccate at the base or not saccate only the inner ones. In B. erucago the petals are yellow, spatulate, of 8-13 mm, and the fruits are asymmetric, indehiscent, 12-20 x 3-6 mm silicles, with four monospermic loculi and with four ribs irregularly toothed or winged at the corners. In B. orientalis the hairs are rounded with claw, yellow, 5-6 mm long and the silicles are 3,5 x 7 mm, ovoidal, covered by small and irregular protuberances, with 1 or 2 monospermic loculi.
Tribe Chorisporeae
It is a tribe with 4 genera and 55 species present mainly in Asia, less in North America and in Europe. They are annuamor perennialplants, usually provided of multicellular glands. The hairs are simple or absent.

The Chorispora purpurascens, native to Iran, Iraq, Israel, Jordan, Lebanon, Syria, Turkey and Arabian Peninsula, belongs to the tribe Chorisporeae formed by 4 genera and 55 species © Ori Fragman-Sapir – www.flora.org.il
The cauline leaves, at times absent, are petiolate and with not auriculate base of the lamina, and with entire or toothed or lobate margins.
The racemes are generally without bracts, usually elongated in the fruit. The flowers are actinomorphic with erect or ascending sepals, and with the lateral couple saccate or not saccate at the base. The petals are white, pink, lavender or violet, rarely yellow with claw, often distinct.
The pollen is 3-colpate and the fruits usually are siliques, with more or less round section, dehiscent or dividing in segments with 1-seed, rarely indehiscent. The ovary contains 5-50, or more, ovules. The style is distinct and the stigma strongly with 2 lobes rarely entire. The seeds are generally uniserial with accumbent cotyledons.
Chorispora purpurascens (Banks and Sol.) Eig is a species native to Iran, Iraq, Israel, Jordan, Lebanon, Syria, Turkey, Arabian Peninsula.
It is an annual plant covered by simple or glandulose hairs, 10-30 cm tall. The lower leaves are petiolate, lanceolate, toothed at the lobes, with a deeply incised lamina. The cauline leaves are similar, but less deeply incised.
The inflorescence is a bractless raceme. The sepals are of about 1 cm, the 1,5-2 cm long petals are violet, with dark veins. The fruit is indehiscent, with a long beak similar to a horn.
Chorispora tenella (Pall.) DC. is a species native to Eurasia but is well known in other parts of the world, particularly in the temperate zones, being a harmful grass. It is an annual herbaceous plant, up to 50 cm tall and abundantly covered by leaves, sticky, due to the presence of periolate glands and of simple hairs.
The leaves are alternate, elliptical-oblong to lanceolate, with 3-8 cm lamina having deeply wawy teeth.The inflorescence is a long raceme. The lower flowers form at the axil of the leaves, whilst the upper ones have no bracts. The calyx is tubular, 6-8 mm, with the lateral sepals saccate at the base. The petals are four, of light magenta or lavender colour.
The fruits are 35-45 mm lomentaceous siliques, curved and ascending, that break under contraints. The upper part of the silique is sterile and forms a 7-20 mm beak. The seeds are subspherical, brown reddish, and get free together with the segment of the fruit.
Tribe Dontostemoneae
The tribe includes 2 genera (Clausia and Dontostemon) with 19 species, mainly present in central Asia with only one species [Clausia aprica (Stephan ex Willd.) Korn.-Trotzky] extending its range in Europe. They are annual or perennial plants, pubescent, with basal leaves arranged in rosette, simple, with entire margins, the cauline, sessile, often alternate, simple. The flowers, regrouped in a terminal raceme, have 4 free sepals with the inner couple lightky saccate at the base, 4 free petals, white or lilac, 6 stamens (tetradynamous) and one unicellular superior ovary. The fuits are dehiscent and linear siliques.

Chorispora tenella, native to Eurasia, is well known as infestant in other parts of the world, particularly in the temperate regions © Ori Fragman-Sapir – www.flora.org.il
Tribe Euclidieae
This tribe includes 28 genera and about 157 species distributed mostly in Asia, and in a lesser number, in Europe, North Africa and North America.
The tribe is characterized by the absence of glands and by the presence of incumbent cotyledons, erect sepals, ramified hairs (rarely simple or absent), entire or bilobate stigmas, often with circular section, with siliques and four-cornered silicles. The base chromosome number is x=7 and, less commonly, x=8.
The results of the analyses of temporal divergence have suggested a Miocene origin of the tribe Euclidieae that probably came from central Asia, from the Pamir high plateau and from western Himalaya. They are annual or perennial plants,rarely shrubs, glandless. The hairs are simple on short peduncle or sessile, but also stellate, biforked, dendritic, rarely absent.

Hesperis pendula, tribe Hesperideae, grows in Jordan, north Israel, Lebanon, Syria and Turkey. Odd brown petals © Ori Fragman-Sapir – www.flora.org.il
The cauline leaves are generally petiolate or sessile, at times subsessile, at times absent, with not auriculate base of the lamina and with generally entire or toothed, rarely lobate, margins. The racemes do not have bracts, and are usually elongated in the fruit. The flowers are actinomorphic with erect sepals, at times ascending up to spread apart. The lateral couple is rarely saccate at the base. The petals are white, pink or violet, sometimes yellow, with claw. The pollen is 3-colpate. The fruits are siliques or silicles, usually dehiscent, not segmented, with circular section or with septa. The ovary contains 2 to 80, at times many, ovules. The style is distinct with entire or strongly bilobate stigma. The seeds are habitually biserial or uniserial axcluding Euclidium. The cotyledons can be accumbent or incumbent.
Tribe Hesperideae
Native to Eurasia, with several endemisms in Greece and in Turkey, this tribe distinguishes for the unicellular glands on one uniserial peduncle. The genus Hesperis counts 50 species whilst Tchihatchewia only one (T. isatidea Boiss.) They are biennial or perennial plants with simple and/or biforked hairs, tardily decentralized siliques, decurrent lobes of the stigma and base chromosome numbers x=5 and x=10.

Diplotaxis acris is an “incertea sedis” species, that is not yet placeable with certainty in a tribe. Present in Egypt, Iraq, Israel and Jordan, grows in rocky but also sandy environments and close to sources. It is edible with known antioxidant properties. Many Brassicaceae are traditionally present in the popular medicine © Ori Fragman-Sapir – www.flora.org.il
The cauline leaves are petiolate, rarely sessile, wiith base of the lamina generally not auriculate and with the margins normally toothed. The racemes do not have bract, long, at times not long, in the fruit. The flowers are actinomorphic with erect sepals with lateral couple usually saccate at the base. The petals are generally white, lavender or violet, with well evident claw. The pollen is 3-colpate. The fruits are siliques, dehiscent, not segmented, with circular section rarely with four corners. The ovary contains 4 to 40, at times many, ovules. The stigma is bilobate. The seeds are uniserial with accumbent cotyledons.
Hesperis laciniata subsp. cupaniana (Guss.) Giardina & Raimondo is a rare endemism of the Madonie that grows on the calcareous gravelly slopes from 1200 to 1800 m of altitude. It is a 15-50 cm tall plant, pubescent-glandulose and with erect-ascending stem, usually not ramified. The basal leaves are spatulate, with toothed margin, the cauline are sessile and with lanceolate lamina. The flowers, arranged in an elongated raceme, have a nastriform corolla, with toothed margins, of violaceous colour. Blooming takes place during the months of May and June. The fruits are cylindrical siliques, supple and velvety.

Striking image of the flowers and of enlarged silique with seeds of Farsetia aegyptia. This desert woody shrub, native to Egypt, can be 50 cm tall, with greyish stems, very ramified, covered by erect and expanded hairs. It is traditionally used by the bedouins for its antidiabetic and antispasmodic properties © Ori Fragman-Sapir – www.flora.org.il
Tribe Shehbazieae
Thanks to the molecular and morphological data now available, this tribe has been created for including the monotypic genus, of hybrid origins, native to Tibet: Shehbazia tibetica (Maxim.) D.A. German, known before 2000 as Nasturtium tibeticum Maxim. It is an alleged hybrid origin of the genus between tribes (Chorisporeae × Dontostemoneae). As a consequence, this tribe presently accomodates the new genus Shehbazia D.A. German, nowadays accepted in the taxonomy of the family Brassicaceae.
They are biennial plants with simple hairs. The root is rather fleshy, the stems are curved, simple. The basal leaves are petiolate and vary from pinnatifid to auriculate. The cauline leaves are like the basal ones, but are little petiolate or sessile, not auriculate at the base. The racemes have no bracts. The lateral sepals are slightly saccate at the base. The petals are white or pink, but appear bicolour bue to tje claw, of intense violet colour. The stigma is deeply bilobate. The ovary contains 12-20 ovules. The fruits are dehiscent siliques, with circular section, straight, with pluricellular verrucae similar to glands. The seeds are spherical and have no wings. The cotyledons are accumbent.

Erysimum pycnophyllum, endemic to Turkey, is used in the popular medicine for treating stomach and lungs diseases, cough, diabetes and compresses when treating hemorrhoids and wounds © Ori Fragman-Sapir – www.flora.org.il
Brassicaceae incertae sedis
Species belonging to different genera such as, for instance, Asperuginoides, Camelinopsis, Chamira, Dipoma, Fourraea,
Idahoa, Lunaria, Paradiodoxa, Subularia do not find a position in any one of th described above tribe, hence they remain excluded waiting for further studies.
Subularia aquatica L. is an annual species diffused in North America and in Europe, known with the common name of water awlwort. This plant grows in ponds, swamps; peat bogs and other shallow cold water bodies, at times on gravel or on sand. It is a small herbaceous plant with 1 to 5 cm long filiform leaves that get thinner at the apex and bears, on peduncles, 2-8 small white flowers. Not all flowers rise above the surface of thr water to be pollinated. Those remaining below the water level, keep close and self-pollinate. The fruits are silicles that free small (less than 1 mm) winged seeds.
The family Brassicaceae counts several species very useful in nutrition and in the human economy.
Seeds of Capsella bursa-pastoris (L.) Medik. have been found in the stomach of the swamp mummy “Man of Tolund”, found in Scandinavia dated of the 300 – 400 BC. The same seeds were found also in the settlings dated 6.000 BC in the important inhabited centre of the Neolithic Era, in the Province of Konya, in Anatolia (Turkey).
It is sure that, since the old times, most of the plants belonging to the Brassicaceae were cultivated and consumed for the human feeding and also for therapeutic uses.
Yore, seeds of Lunaria annua were employed as diuretics and the whole plant against scurvy and as digestive. Recent researches have proven that the plant contains, among the active principles, fatty acids, enzymes, glucosides, minerals, vitamin C and the alkaloid lunarina having a strong sour taste. The leaves are edible, their taste recalls that of the cabbage, but more bitter. Also the flowers and the unripe fruits are edible and can be added to mixed salads and omelettes with broiled eggs.
Diplotaxis acris (Forssk.) Boiss is a species diffused in North Africa (Egypt), Arabian Peninsula, and western Asia (Iraq, Israel and Jordan). It is an annual herbaceous plant, 10 to 50 cm tall, that grows in rocky, but also sandy, environments and close to water sources.

The white mustard (Sinapis alba subsp. alba) has digestive, carminative, anti-inflammatory and stimulating properties. External compresses of leaves against rheumatisms © Dr. Giuseppe Mazza
The basal leaves, with jagged margins, are juicy and are arranged in rosette. If consumed in salad, they have a peculiar spicy flavour similar to that of the watercress, and for this reason, it is called “garden rocket”. It is eaten also by animals such as goats, sheep and camels. The flowers, in dense racemes, are of violet colour and the fruits are erect siliques. Recent studies have evidenced its antioxidant properties.
Erysimum pycnophyllum J. Gay (= Erysimum thyrsoideum Boiss.) is a species endemic to Turkey, traditionally used in the popular medicine for treating stomach and lung disorders, cough, diabetes, and used in compresses on the piles and on the wounds. Phytochemical studies have emphasized the presence of alkaloids, anthraquinones, coumarins, flavonoids, saponins, tannins, glucosides and essential oils.
A species native in Europe and in western Asia, is Cardamine pratensis L. so called being peculiar the the humid meadows on acidic soils in the water meadows, from 0 to 1300 m of altitude.

The leaves of Nasturtium officinale are employed as diuretic for the kidney problems © Dr. Giuseppe Mazza
In Italy, its leaves are consumed raw, in salads, alone or with other raw vegetables, or with boiled potatoes, cooked in the vegetable soups. In phytotherapy, the plant is employed for its tonic, antianemic, mineralizing and antirheumatic properties.
In Bolivia the herbal therapy is practiced by the local healers since hundreds of years and at least four are the species employed for this purpose. The leaves of Capsella bursa pastoris L. are utilized in infusion for kidney problems. The infusion of leaves of Lepidium bipinnatifidum Desv. is employed in the gastro-intestinal disorders and diarrhoea, and, in a reduced concentration, in the children as tranquillizer. The leaves of Nasturtium officinale W.T. Aiton are employed as diuretic for the kidney problems. With the leaves of Sinapis alba subsp. alba L. (=Brassica hirta Moench) are prepared external compresses against rheumatisms.
Lepidium draba L. is a perennial herbaceous plant, native to the Mediterranean arid zones and of Central Asia, dangerous for the cultivations as it is attacked by various viral parasites and by various species of insects and of mites that in turn spread in the cultivated species. In the past its leaves were utilized for its antiscorbutic and carminative properties and also as seasoning for food.

Cochlearia officinalis is a plant rich in vitamin C, with purifying and stomachic properties © A.S. Kers
In Europe, the shepherd’s purse (Capsella bursa-pastoris) is utilized for its strong anti-haemorrhagic and regulating properties of the menstrual cycle.
Cochlearia officinalis L. since the XVI century has been used as plant capable to cure the disease known as scurvy, caused by the deficiency of ascorbic acid (vitamin C) in the body. It also has other properties, and has been utilized in phytotherapy as purifier, detergent, rubescent and stomachic.
Farsetia aegyptia Turra is a perennial woody shrub native to Egypt. It mostly grows in rocky places and in presence of little deep sand. It has 15-20 cm tall stems, greyish, covered by erect and extended hairs. The bedouins utilize this plant for its antidiabetic and antispasmodic properties.
In Japan, the Japanese horseradish (Eutrema japonicum (Miq.) Koidz.), known as wasabi, has gained its specific pungent flavour during its long history of domestication and from it have derived several varieties.

The turnip (Brassica rapa) is widely consumed as a vegetable. Varieties with leaves, root, cymes and oil seeds © Dr. Giuseppe Mazza
The most diffused are E. japonicum var. Daruma and E. japonicum var. Mazuma. They are utilized utilized for the leaves as well as for the rhizome: the first ones, dried up, serve to flavour dishes; conversely, the second is used for seasoning the green paste, also called wasabi, to accompany raw fishes, ln particular, the sushi and the sashimi. In the sushi is smeared a small quantity of it between the fish and the rice, and in the sashimi it is mixed in small quantities in the soy sauce where the raw fish is to be dipped . This use has become a peculiarity of the Japanese cuisine ad culture.
Brassica oleracea L. stands among the most important species due to the diffusion and the alimentary use of the leeaves, of the inflorescences, of the flowered heads, of the buds and of the roots. Whilst Brassica rapa L. is the ancestor of all the many varieties, cultivars, local ecotypes of turnips, most of them with a biennial reproductive cycle.
The numerous varieties of cabbage owe their origin to morphological characteristics fruit of the crosses between plants having mutations affecting very few genes the man has maintained in the time thanks to their great capacity of environmental adaptation.
The process of deomestication that has resulted in the cultivated forms of the cabbage is not yet completely clear, neither for what concerns its initial localization, nor for the progenitor species involved. We can suppose that the man has done a disruptive selection on the spontaneous plants favouring the develoment of some of their organs. By selection of the terminal buds has originated the cabbage, by selection of the lateral buds have taken form the Brussels sprouts, by selection of the stems has been obtained the kohlrabi, by selection of the leaves was gotten the curly kale, by selection of the stems and the flowers, has formed the broccoli and by selection of the inflorescences the cauliflower.
The possible wild progenitors are to be searched in the genus Brassica and their genome, as assumed by Nagaharu U, is of type CC and base chromosome number 2n=18.
About the range of origin of the cauliflower, two alternative hypotheses suggest two locations: one in north-western Europe and one in the Mediterranean area.
The studies of molecular genetics and, in particular, the study of the polymorphisms of the length of the fragments of nuclear restriction RFLS (Restriction Fragment Length Polymorphism) have found that the B. oleracea subsp. oleracea is the closest to the cultivated species, and, therefore more strictly related to the ancestors of all cultivated B. oleracea, whilst the other wild species of Brassica, with Mediterranean distribution, result being more distantly related.

Brassica oleracea var. botrytis is usually known in this way. It’s the classic white and roundish cauliflower present in all kitchens © Dr. Giuseppe Mazza
As B. oleracea subsp. oleracea is found only on the Atlantic coasts of western Europe, in Great Britain, France, northern Spain and on the islands of Helgoland and of Rügen (Germany), it has been advanced the hypothesis that the domestication had happened on the Atlantic coast and from there introduced in the Mediterranean, where the following selection took place. However, othe Authors believe that the Mediterranean domestication, in particular of the eastern area, is supported by linguistic and literary considerations, as witnessed by the Greek and Latin literatures, confirming, since the origins, the knowledge and the use of the cultivations of B. oleracea. Whilst no other historical document indicates the use of these plants in the Fertile Crescent and in the old Egyptian civilizations.
It is well known that some forms of B. oleracea ssp. oleracea were cultivated in Greece already in the IV century BC, whilst the diversification of the cultivation with other types has begun later. Theophrastus (IV-III BC) described three types of brassica, one of which wild, characterized by smooth leaves, acrid taste and utilized for medicinal purposes; the other two were cultivated and differed for the crunchy leaves in one type and for the smooth leaves and reproductive difficulties in othe other one. This last type may be related to one of the old initial forms of cauliflower with a lower fertility of the flowers.

Romanesco, also known as Romanesco broccoli or Roman cauliflower, is a particular cultivar of Brassica oleracea var. botrytis. Similar to cauliflower differs for the pale green colour and in the pyramidal structure with many rosettes. The smaller ones, spirally arranged, are repeated, decreasing until they merge, with some regularity © Dr. Giuseppe Mazza
Cato the Elder (234-149 BC) used the term brassica and praised its health properties. Also Pliny the Elder (23-79) mentioned some types of brassica cultivated in Roman times, especially in the Italian Peninsula, describing its characteristics very similar to the cauliflower and to the broccolis. Latin texts of the IV century AD describe the morphology, the agricultural practices and the relevant culinary and medicinal uses of these brassicas offering, inter alia, a valuable contribution to the ethnobotanical knowledge of those peoples.
In the Middle Ages, the juice of cabbage was mixed to honey for treating the hoarseness, cough and all lung diseases. It is possible, therefore, that the cauliflower has got origin direcly from the broccolis in the western Mediterranean basin.
It is likely that it had been introduced in the Italian Peninsula from Cyprus around 1490, thanks to the Genoese and Venetian navigators, and then carried to France, Belgium, Great Britain during the XVI century with the name of “Cyprus cabbage turnip” as stated by the Flemish botanist and physician Rembert Dodoens (1517-1585), who in his opus Cruijdeboeck, published in 1594, represented and called under this name the plant that only later on was called “cauliflower”. It spread throughout Europe mostly during the XVII and XVIII centuries and then all over the world.

The Savoy cabbage (Brassica oleracea var. sabauda) has wrinkled, crinkled leaves with prominent veins. It is usually utilized for soups and broths © Dr. Giuseppe Mazza
As confirmation of the data reported by the past literature, recent studies of molecular biology indicate that the species endemic to the Aegean Brassica cretica Lam. is the extant closest relative to the cultivated B. oleracea. It is consequently confirmed, that the domestication of the cultivated brassicas occurred in the eastern Mediterranean region.
The vegetables belonging to the Brassicaceae are consumed everywhere in the human diet and are considered as very important. in Europe and also in China, Japan, India. It is estimated that in all the world are consumed mainly cauliflowers, cabbages, broccolis, turnips and kohlrabis also due the presence of various substances useful for the health, such as carotenoids, ascorbic acid and the phenolic compounds that can control various diseases related to cancer, heart and the degenerative ones. Recently, it has been discovered that the indole-3-carbinol (I3C), substance derived from the degradation of the glucobrassicin (glucosinolate present in the cabbages, broccolis, cauliflowers and Brussels sprouts) is able to block the replication in the cells of the coronavirus SARS-CoV-2, hindering its spread in the rest of the organism. The results have been obtained in laboratory, and pending clinical trials on man, they are quite confident that this substance can be very useful to the subjects already infected by COVID-19.

The broccoli, or broccoli cabbage (Brassica oleracea var. italica) has ramified globular inflorescences, generally of green colour but at times red or violet, and several big costoluto leaves. In kitchen, besides being used in preparing soups and broths, is employed steamed, au gratin or stir-fried, or dipped raw in pinzimonio © Dr. Giuseppe Mazza
Many are the known and utilized varieties of B. oleracea, we here mention the most important ones.
The black cabbage, also called cabbage knight, (B. oleracea var. viridis L. = B. oleracea var. acephala) is a variety employed in the Nordic countries for feeding the livestock. It has long and bullate leaves, of dark green grey colour and in Italy the leaves, stemless, are employed in the typical Tuscan recipes (ribollita, bread soup and farinata).
The Chinese broccoli or [B. oleracea var. alboglabra (L.H. Bailey) Sun] has thick leaves, smooth and of green-bluish colour with large petioles and inflorescence similar to that of the broccolis, but smaller, with just a little more bitter taste. In the Chinese cuisine it is sautéed with ginger and garlic, or steam cooked to be served with an oysters sauce.
The kale or leaf cabbage (B. oleracea var. sabellica L.) is very diffused in Germany and also in other north-European countries and on the eastern coast of North America.

The Brussels sprouts (Brassica oleracea var. gemmifera) grown mostly in northern Europe. Edible axillary buds, rich in vitamins, fibres and glucosinolates very active against lung and digestive system cancers. They are consumed usually steamed with some butter, at times with addition of lemon © Dr. Giuseppe Mazza
In the kitchen it is used for light or thick soups, at times also for the stew, with other vegetables, or as side dish. The leaf cabbage, with potatoes, smoked sausage, fat bacon, salt, pepper and some milk, is an ingredient of the traditional Dutch dish called boerebkoolstamppot.
The Savoy cabbage or cabbage (B. oleracea var. sabauda L.) is a variety resembling the white cabbage, but differing from this for the presence of wrinkled, rippled leaves, with prominent veins. It has very ancient origins, and nowadays is diffused in most of Europe and in Italy,in the central-northern regions. In the kitchen it is employed for preparing light and/or thick soups. In the Piacenza region, the Savoy cabbage is appreciated for the risotto with pork ribs.
The broccoli, or broccoli cabbage, (B. oleracea var. italica Plenck) has inflorescences generally of green colour, but at times red or violet, differing from those of the cauliflower because globular and ramified. The leaves are numerous and costolute. It is diffused in most of Europe and of the world. In kitchen, it serves for preparing light and/or thick soups. It can be consumed also raw with pinzimonio, but in various regional recipes it is employed steam cooked, au gratin or stir-fried.
The cabbage (B. oleracea var. capitata) differs from the Savoy cabbage due to its smooth leaves, arranged in form of a compact ball. In the kitchen it can be consumed raw, very finely cut, and served in salad, or employed for preparing the sauerkrauts that are eaten, generally boiled, as side dish of the pork shank, of the black puddings and of other types of sausage. Sauerkrauts are obtained by fermentation of the leaves cut into strips placed in layers in wooden casks, with salt, pepper and spices. In the western world, were the Greeks and the Romans to ferment the cabbages whilst in Asia it was already a centuries-old habit to utilize the lactic fermentation to preserve the vegetables. There is also the purple cabbage that is excellent if consued raw, in salad.
The red cabbage (B. oleracea var. capitata L. f. rubra) is also known as violet cabbage or ref or blue sauerkraut and has dark red leaves, tending to violaceous. The red cabbage, very resistant to cold, is diffused in northern Europe, in America and in China. It may be eaten raw, alone or as an appetizer in mixed salads, to accompany different dishes, such as, for instance, the sauerbraten, a type of German stew marinated and then cooked in water, vinegar and spices.
The Brussels sprouts (B. oleracea var. gemmifera DC.) are a variety with axillar edible buds and are grown mostly in northern Europe because they adapt effectively to the cold climate. They are eaten steamed with a little butter, sometimes with addiction of lemon. They are rich of vitamin B6, vitamin C, fibres and potassium as well as of glucosinolates (glycosidic compounds containing sulphur), very active against the cancers of the lung and of the digestive system.
The cabbage turnip (B. oleracea var. gongylodes L.) is native to western Siberia. The lower part of the stem (called “stalk”) forms a spherical swelling over the soil of fleshy consistency and is suitable for numerous preparations.

The cabbage turnip (Brassica oleracea var. gongylodes) has stem big and fleshy, globoso, with white and crunchy pulp. It can be eaten, with or without leaves, cooked or raw in salad. © Dr. Giuseppe Mazza
Also the leaves are edible and have an excellent sweet and cool taste. The globose form has red or violet or green shades and white and crunchy pulp. It can be consumed either cooked or raw in salad.
The cauliflower (B. oleracea var. botrytis L.), whose name comes from the Latin caulis (cabbage) and floris (flower), is formed by a big corymbous inflorescence, called head or curd, built by numerous enlarged and thickened floral peduncles and that represents the edible part of the vegetable.
The curd can assume, depending in the various cultivars, a different colouration (white, straw-coloured, green, violet). The cauliflower is consumed boiled, fried, roasted, steam cooked or raw. It may be used as side dish, in salads or as main course in the vegetables soups.
The Romanesco broccoli, also called Roman cauliflower or Romanesque cauliflower, is a particular cultivar of the cauliflower. It is very similar in form, but differs in the colour, that is pale green, and in the structure, that is pyramidal, with many rosettes, in turn composed by other smaller rosettes, spirally arranged, that repeat with a certain regularity.
In the kitchen it is prepared boiled, but also stir-fried. The Romanesco broccoli has a more aromatic flavour than the broccoli, contains more vitamins and minerals.
The Portuguese cabbage (B. oleracea var. costata DC.) is an up to 90 cm tall plant, with fleshy and succulent leaves utilized in the soup known as caldo verde (literally, “green broth”), typical to Portugal and Galicia but very diffused and utilized also in the Portuguese-speaking countries and, in particular, in Brazil. It is a simple dish that can be served as appetizer or as second course of soup. It has a green colouration because the cabbage is cut in very thin strips to which are added potatoes, onions, garlic and spicy sausages. The Galicia cabbage is also an important plant in the production of honey.
The twiggy turnip (B. fruticulosa Cirillo), in Sicily called “cavolicello”, is an annual plant that in nature is present in some zones of central-southern of Italy. It is consumed cooked, simply boiled and seasoned with oil, or in pan with garlic and chili pepper or stewed with wine. It has a pleasantly bitter taste and in Sicily, is utilized as side dish for the grilled sausage.
The rapeseed (Brassica napus) did form about 10.000 years ago by hybridization and originates from the fusion of two genomes, that of B. oleracea (with genome CC, base number x=9) and that of B. rapa (with genome AA, base number x=10) and has genome AACC, base number x=19. The cultivation has begun in India in 4.000 BC, in China and in Japan 2.000 years ago and in Europe during the Middle Ages and then has diffused all over the world for the utilization of the seeds very rich of oil.

Section of red cabbage (Brassica oleracea var. capitata f. rubra) with the typical dark red leaves. Can be eaten raw, alone or as appetizer in mixed salads, or cooked as side dish for various preparations, such as the Sauerbraten, German marinated beef stew and then cooked in water, vinegar and spices © G. Mazza
The mustard [B. nigra (L.) W.D.J. Koch] is cited by Jesus in one of His parables: “The kingdom of heaven is like a mustard seed, which a man took and planted in his field. Though it is the smallest of all seeds, yet when it grows, it is the largest of garden plants and becomes a tree, so that the birds come and perch in its branches.” (Mt. 13,31-32; Mc. 4,30-32; Lc. 13,18-19). Known also as black mustard, its is native to part of Europe, of the Mediterranean region, and to other parts of North Africa and has been naturalized in Great Britain, in North America and in New Zealand.
The consumption of seeds of black mustard is useful to improve the mechanisms of immune defence against cancer. Specific studies have proven that it can reduce the rate of carcinogenesis of the colon, bladder and lung. It has antioxidant activity and the presence of polyphenol inhibits the enzyme α-pancreactic amylase resulting in the improvement of the glucose tolerance in the diabetics. It is also used against headaches and neuralgias.
The turnip (B. rapa L.) has its ancestors in the area of the so-called Fertile Crescent corresponding to the regions of the highlands in the present Iran, Iraq and Turkey. The first human hunters-collectors and farmers discovered that the leaves and roots furnished food and had medicinal properties and did consume them either raw or cooked. In the times of ancient Greeks and Romans, the turnip was consumed only by the plebeians. In the Middle Ages and untile the XVIII century, it was utilized as winter food for the animals.
The Europeans introduced it in USA, in Canada, in Australia and in New Zealand and now is cultivated in the whole world as vegetable, of which are eaten, in the different varieties, the leaves, the fleshy root (turnip), the flowered tops, the oily seed. The turnips are little calorific, but are rich of mineral salts (potassium, phodphorus, calcium and sodium), vitamin C and amino acids.
The seeds are very nourishing because of the high contents of vitamins (A, B1, B2 and C) and of proteins. In the Australian phytotherapy its use has been documented as liniment to relieve muscle pain, to disinfect wounds and in the patches for the remedy of chest congestion and bronchitis. The tribes of central Himalaya utilize B. rapa subsp. oleifera (DC.) Metzg. for treating various injuries in the animals such as bone fractures, burns and accidental cuts.

Cultivar of Brassica oleracea var. botrytis with the head of various colours. Like the white ones, they are eaten boiled, fried, roasted, steamed or raw © Dr. Giuseppe Mazza
The napa cabbage or Chinese cabbage [B. rapa subsp. pekinensis (Lour.) Hanelt] has a good taste and is the main ingedient of the Korean kimchi formed by fermented vegetables salted or seasoned for seasoning, but can be served as main course. It is also used in the preparation of other Korean dishes such as the kimchi jjigae and the kimchi bokkeumbap (pan-fried rice with meat or fish and vegables rolls). It can be stir-fried with other ingredients such as tofu, mushrooms and zucchini. The kimchi is part of the Korean culture and identity and, for such reason, has been inserted by the UNESCO in the list of the Intangible Cultural Heritage of the Humanity.
The rapini or broccoli rabe [B. rapa L. subsp. sylvestris (l.) Janch. var. esculenta Hort.] is a plant of Mediterranean origin and differs from the common turnip for the annual cycle and the taproot root that does not get bigger. It is a vegetable typical of the Italian agriculture mainly cultivated in Puglia, Campania, Molise, Lazio and Calabria for the utilization of the inflorescences and of the tender part of the stem with the leaves. It is consumed in the typical dishes of the Apulian cuisine, such as the orecchiette with the rapini, the strascinati with rapini, stewed turnips with chili.

The root of black radish (Raphanus sativus var. niger) is traditionally used to stimulate biliary function © G. Mazza
In central and in northern Europe, as well as in Italy, in the Po Valley, the swede, that is an annual or biennial cultivar of B. rapa, and the rapeseed, (B. napus) are cultivated as forage crop and also for oil cakes, after the squeezing of the seeds for a feeding of the livestock rich in proteins.
The calepina [Calepina irregularis (Asso) Thell.] is native to the countries of central Asia and of the Mediterranean Basin and naturalized in central Europe, North America and Australia.
The leaves can be consumed as vegetable but its use in the kitchen is totally unknown. It is rich of vitamins and mineral salts and has remarkable diuretic and detoxifying effects.
The white mustard (Sinapis alba subsp. alba L.) has digestive, carminative, anti-inflammatory and stimulating properties. In the past, in order to get a good anti-inflammatory effect, they prepared doughs of seed flour and water (sinapisms) to be locally applied. The poultices were employed also for treating the rheumatisms. With the green plant they realized a decoction used as vasodilatory and added to the water for the footbaths.
The toasted seeds of white mustard are employed in various baked goods, if chopped for preparing omelettes, soups and desserts.
The species Cardamine diphylla (Michx.) Alph.Wood, native to Canada and to the central-eastern USA, is employed by the Iroquois as infusion to strengthen the breasts. They also use to chew the raw root for the gastric gasses and make root poultices on the swellings. The cold infusion is utilized to reduce the fever and the beginning of the tuberculosis. The raw roots with salt or boiled entire are employed in the kitchen.
The horseradish (Armoracia rusticana G. Gaertn., B. Mey. & Scherb.), presumably native to south-eastern Europe and western Asia, is cultivated since more than 2.000 years for its roots and, as the cultivated plants are generally sterile, is propagated planting pieces of root like many cultivations propagated vegetatively.
When entire, the root of horseradish has no aroma, but, if cut or grated, some enzymes do activate releasing its peculiar spicy flavour. With the horseradish they prepare a spicy sauce that goes well with boiled meats, smoked fish and many other dishes.
The hoary mustard [Hirschfeldia incana (L.) Lagr.-Foss.] is native to the Mediterranean Basin but has been diffused in many parts of the world often as noxious weed. It has lobate leaves that form an ample basal rosette.

The rocket (Eruca vesicaria subsp. sativa) is an annual herbaceous plant known since the old times © G. Mazza
This species looks a lot like the black mustard, but differs because it is perennial and smaller. The stem and the leaves are covered by soft white hairs. Its leaves are edible and in the past they were used as leaf vegetables. Still today, in Sicily, they are utilized in gastronomy as a sauce for pasta and meats thanks to its intense and somewhat bitter taste.
The rocket [Eruca vesicaria subsp. sativa (Mill.) Thell.] is an annual herbaceous plant known since the old times, native to the Mediterranean Basin, and central-western Asia, presently cultivated in North Europe, in North America and in India.
In the kitchen are utilized the leaves when still tender and before the development of the floriferous scape and are consumed fresh in salads, or to garnish cold cuts or raw slices of meat or fish or on top of pizzas and focaccias after cooking. With the leaves of rocket may be prepared an aromatic pesto with which to dress pasta and bruschettas.
The Romans utilized it, and consumed even its seeds, for its medicinal properties and particularly for its aphrodisiac properties as well as for its antiscorbutic and antibacterial functions.
The rocket is grown on a large scale in China for the extraction of the oil from the seeds, because rich of erucic acid, important in industry, but also for lighting and as lubricant. The young plants are employed for medicinal purposes, as stimulant, antiscorbutic, gastric and diuretic.
Another species of rocket, that grows spontaneous in central-southern Europe, is Diplotaxis tenuifolia (L.) DC., known as wild rocket or perennial wall-rocket distinguishable from the grown one by the more serrated leaves, the yellow flowers (the cultivated rocket normally has white flowers or of straw colour) and a much stronger and spicier flavour. It can be utilized raw for flavouring salads or in the same way as the cultivated rocket for pastas and fillings.
The radish (Raphanus sativus var. sativus L.) is a plant that can be one metre tall and that presents an enlarged taproot root normally of bright red colour (but many varieties with different colours do exist).
The basal leaves are petiolate, lirate and divided in segments, the cauline ones are entire, of lanceolate or oval shape and toothed margins.
The root of black radish (R. sativus L. var. niger) has been utilized in the popular medicine since ancient times as natural medicine for stimulating the biliary function. Specific studies on laboratory rats, have confirmed that the juice of the root has significant antioxidant properties with a beneficial effect in the alimentary hyperlipidemia.

Rapeseed (Brassica napus) has formed by hybridization of two genomes, that of B. oleracea and that of B. rapa. Its cultivation started in India 4000 years BC, in China and in Japan 2000 years ago, and Europe during the Middle Ages © Dr. Giuseppe Mazza
The wild radish (R. raphanistrum L.) is a 20-80 cm tall herbaceous plant native to Mediterranean, now present also in south-western Asia, in southern Africa and in eastern coastal North America. It has 8-15 cm long leaves, pinnato-lirate with 5 or more segments, petiolate, hairy-shaggy with toothed margins. The cauline leaves are progressively reduced and sub-sessile. The inflorescences are terminal racemes formed by several flowers supported by short pedicels. The calyx is violaceous and the corolla has the spatulate petals, alternated to the sepals. The ovary is superior bicarpellate surmounted by a style having a capiate stigma. The fruit is a lomentaceous silique, pointed at the apex, pedunculate, not dehiscent.
The radish is known since several millennia as witnessed by the archaelogical finds from the ancient civilizations of Greece, China and Egypt. After Pliny the Elder, the Greek physician Androcydes (IV century BC – III century BC) recommended their use to Alexander the Great in order to avoid the harmful consequences genereted by the excessive use he made of the wine. In various parts of the world the leaves and the roots of this plant have been utilized to treat cancer and as antimicrobial and antiviral agents.
The cultivations of oily seeds of Brassica, with the 14% of the edible vegetable oil in the world, represent the third source after the soybean [Glycine max (L.) Merr.] and the oil palm (Elaeis guineensis Jacq.).
The Soviet agronomist Georgii Dmitrievich Karpechenko (1899 –1941), specialized in vegetable cytology has created various hybrids crossing the radish (Raphanus sativus) with the cabbage (Brassica oleracea), both with 2n = 18 chromosomes. In the first generation he has obtained only sterile hybrids with a diploid chromosomic set. Working experimentally, he found that the tetraploid hybrid (2n=36) that combined the whole diploid complement of the radish as well as that of the cabbage, could undergo a normal meiosis proving to be fertile, morphologically distinct and reproductively isolated, from both parental species. Karpechenko believed having found experimentally found a “new species” that he called Raphanobrassica as it combined the genetic material of two extant genera. These discoveries were announced by himself in 1927 in Berlin during the Fifth International Congress of genetics, followed by publications in Russian (1927) and in English (1928). His researches, unfortunately, were interrupted by the agents of the People’s Commissariat for Internal Affairs who indicted him with the false motivation of belonging to an alleged “anti-Soviet group” and sentenced him to death executing him on July 28th, 1941.

The rapeseed is now diffused all over the world due to the seeds quite rich in oil. After squeezing, the compact residue is reduced in oil cakes and used as feed for livestock © Dr. Giuseppe Mazza
Presently are known two other fertile forms of Raphanobrassica. One is the allopolyploid hybrid x Brassicoraphanus (2n=38), obtained from the crossing between Raphanus sativus and Brassica rapa, that is cultivated as fodder plant and Raphanofortii (2n = 38), allopolyploid hybrid between Brassica tournefortii (2n = 20) and Raphanus caudatus (2n=18).
The changes of the society from the rural to the industrial stage, with consequent high urbanization and abandonment of the countryside have favoured the work in the factories. This industrialization resulted in the release into the biosphere of a dramatic quantity of polluants that worry increasingly and require great global attention.
The extractions, the metal castings, the utilization of pesticides and the industrial discharges are the main sources of pollution.
It has been proved that most genera of the family Brassicaceae have the capacity to accumulate various contaminants and are therefore in condition of cleaning up the environment where they do grow.

Plant of radish in bloom (Raphanus sativus var. sativus). The enlarged taproot, with a lightly spicy taste, has white or scarlet skin. It is consumed raw, and sliced or artfully carved it ifet decorates the serving dishes © G. Mazza
The genera Alyssum, Arabidopsis, Brassica, Odontarrhena and Thlaspi are able to have an overaccumulation of metals and hence play a key rôle in the technology of the phytoremediation.
The overaccumulation of Ni is a rare form of physiological specialization shared by plants that often grow in the serpentines. Most of these species take advantage from the high levels of this metal naturally present in the ultramafic rocks, accumulating it as defence against the herbivores.
Many are the endemic species living in the serpentine soils (ecological endemisms of the sepentine) ans accumulate Ni at high concentration. A high accumulation of Cr is rare or never adequately demonstrated.
However, not all species are utilizable for the environmental reclamation (bioremediation), especially if they have a slow growth and a low biomass yield.
Conversely, Brassica juncea, native to Asia and diffused in central and southern Europe and in North America, is a very interesting species for the bioremediation, because it is able to accumulate 34,5 mg/kg of Pb in the buds and has a high growth in biomass though its optimal growth is limited to the low values of pH and to high concentrations of metal in the soil. It is able to absorb also, selenium, sulphur, chrome, lead, copper, cadmium, nickel and zinc.
It is utilized also as green manure plant because it allows to control, thanks to the high contents of glucosinates, nematodes and pathogenic fungi present in the soil such as Sclerotinia, Rhizoctonia, Pythium. It is also utilized Odontarrhena oxycarpa (Boiss. & Balansa) Španiel & al. due to its excellent capacity of accumulating Ni with concentrations reaching the threshold criterium of 1000 mg/kg.
Noccaea caerulescens (J. Presl & C. Presl) F.K. Mey. (=Thlaspi caerulescens J. Presl & C. Presl) is an extremophile plant capable of overaccumulation of heavy metals, such as zinc, cadmium and nickel.
The family Brassicaceae includes also ornamental plants that are found in the genera Aethionema, Alyssum, Arabis,
Aubrieta, Aurinia, Cardamine, Erysimum, Heliophila, Hesperis, Iberis, Lobularia, Matthiola and Lunaria.
A plant that can be utilized for adorning sunny rocky gardens in the cold regions is the Persian stonecress (Aethionema grandiflorum Boiss. & Hohen) native to eastern Turkey, northern Iran and Caucasus. It is a shrubby perennial plant forming spots of morbid blue-green needle-shaped leaves and during the summer months covers of pink flowers. It grows well in poor, dry and well drained soils and does not love humidity. After blooming it can be pruned in order to keep a bushy posture.
Cardamine pratensis is an ornamental plant cultivated in the European gardens, but also introduced and naturalized in North America.

Aubrieta deltoidea is cultivated as an ornamental plant in the rocky gardens. Small leaves and abundant pink, violet or blue flowers from spring till late summer © Ori Fragman-Sapir – www.flora.org.il
Odontarrhena oxycarpa is a small compact plant coming from Turkey with greyish leaves and racemes of yellow flowers, ideal to be growing in the cracks.
Iberis sempervirens L. is a species suitable for rocky gardens but can also be employed in the open ground to form spots or borders and even in pots. It has 1 cm leaves, long, lightly coriaceous and persistent whose colour varies depending on the temperature. In the winter months they are more glossy and have a bright green colour. The flowers are small and are of candid white colour united in racemes covering all the plant. It is a plant of easy cultivation, fairly rustic and does not displat pathologies or parasitic attacks.
Lunaria annua is a biennial, or perennial and rareky annual herbaceous plant. It is commonly called Pope’s money, coins of Judas, perennial honesty, silver dollars, Chinese money. It has cylindrical stems, erect, hairy and ramified that may reach the height of 1,20 m. The leaves, of bright green colour, are heart-shaped, deciduous, opposite with toothed margins.

Erysimum cheiri has several fragrant flowers. Green-violaceous sepals and petals that, in cultivars, have yellow, orange, red, bordeau, violet, brown and cream shades © Dr. Giuseppe Mazza
From March to May, it produces apical racemes of flowers weakly perfumed, formed by four purple, or even white, petals. The beauty of the plant is also given by the fruits that are erect or a little hanging silicles, rather large (up to 6 cm), dehiscent, flattened, subrotund, of greenish colour where not yet ripe, silvery with two valves and with the replum, when ripe.
Cardamine pentaphyllos (L.) Crantz is a species very utilized as ornamental plant. Because of the beauty of its leaves, of its capacity of growth, for the stability of its forms, for the colour of the flowers and for the resistance to parasites, it has got the prize “Award of Garden Merit” of the Royal Horticultural Society.
C. pratensis is cultivated in the European gardens, but is also introduced and naturalized in North America.
Very ornamental is Aubrieta deltoidea (L.) DC., native to south-eastern Europe, cultivated in all the world as ornamental plant in the rocky gardens, on dry stone walls, where it creates suggestive cushions of small roundish leaves, grey-green, and produces, from spring to late summer, abundant small flowers of pink, violet or blue colour.

Due to its ornamental characteristics Cardamine pentaphyllos has got the prize “Award of Garden Merit” of the Royal Horticultural Society © Dr. Giuseppe Mazza
Matthiola incana (L.) W.T. Aiton, native to the Mediterranean regions, is a 30-60 cm tall plant, known as red violacciocca, suitable to be cultivated in open ground as ornament of gardens, or in pots, or for its cut flowers. Several varieties of it do exist, mostly annual, with simple or double flowers of various colours (white, pink, red or violet).
Hesperis pendula DC. is a perennial herbaceous, 30-60 cm tall plant growing in Jordan, northern Israel, Lebanon, Syria and Turkey. It’s sticky and hairy, with short glandular hairs and white hairs with rigid bristles. The stems are erect or prostrate, green, lightly ramified towards the apex. The cauline leaves are small. The inflorescence is a raceme that elongates in the fruit. Blooming takes place by end April and early May and the flowers present the corolla with four yellowish petals, brown when ripe. The fruits are hanging siliques, thin, 8-5 cm long, covered by glandular hairs and hairs dimple and ramified.
Hesperis matronalis L. is a plant with whoch the Roman women used to adorn themselves with its flowers during the Matronalia, feasts that were celebrated during the celends of March in memory of the the “Sabine women rape”.

Hesperis matronalis, so called because the Roman women carried this flowered plant, together with incense, to the temple of Juno Lucina during the Matronalia, festival celebrated the March calends, in memory of their work of peace after the “Rape of the Sabine”. Fragrant flowers on dense racemes that elongate after the anthesis © Jean Louis Verbruggen
In fact, it is known as dame’s violet and is employed in the gardens for the beauty of its flowers, very fragrant, carrfied by dense racemes that elongate after the anthesis. The name of the genus Hesperis probably refers to the scent of the flowers that gets more evident by the evening (from the Greek hespérios, vespertine). Two subspecies are present in Italy: H. matronalis L. subsp. matronalis and H. matronalis subsp. nivea (Baumg.) E.P. Perrier.
As well as being an ornamental plant it is utilized in the traditional medicine for its stimulant, antidiarrheal, sudoriferous, diuretic and expectorant properties. The leaves have also rubefacient properties and, in the past, they were used in poultices for favouring the maturing of the abscesses.
Erysimum cheiri (L.) Crantz (=E. helveticum (Jacq.) DC.), known as wallflower, is a species native to the Aegean and later on introduced in all Europe as ornamental plant due to its numerous flowers with green-violaceous sepals and round petals two to three cm long, of bright yellow, red, or violet colour that form in spring and are very fragrant. Many cultivars do exist, in yellow, orange, red, burgundy, violet, brown, white and cream shades.

Odontarrhena oxycarpa, native to Turkey, is used as ornamental plant and for the remediation of the polluted sites due to its excellent capacity of storing nickel © G. Mazza
In the past this plant was utilized for its medicinal properties because believed to be aphrodisiac, antirheumatic, antispasmodic, cardiotonic, diuretic, emmenagogue, expectorant, ophthalmic, purgative, stomachic and tonic.
E. rhaeticum (Schleich. ex Hornem.) DC., species known as “Swiss wallflower” or “Rhaetian wallflower”, is endemic to central-western Alps and nearby areas, grows in the arid meadows, on stony slopes, in the brushwoods up to 1800 metres of altitude and blooms from April to August.
Heliophila coronopifolia L. is an annual species endemic to southern Africa, up to 70-80 cm tall, a little hairy in the basal parts. The leaves, often stipulate, are pinnatisect. The flowers, of blue colour with white or greenish centre, open from August to October. This species, easy to cultivate, is utilized as ornamental plant in the garden. It grows quickly if placed on well drained soil, in full sun, and with regular waterings during the growth season.

Aethionema grandiflorum is a shrubby perennial plant with needle-shaped leaves. In summer it adorns the sunny rocky gardens of the cold regions © Paolo Gislimberti
The process of selection done by the man has also produced ornamental cabbages up to 30 cm tall and up to 20 cm broad. The leaves are big, with toothed margin, of green and white, green and pink, green and violet, colour.
The ornamental cabbages are very appreciated also for the leaves coloured with very showy dyes ans are able to resist very well to cold and frost.
Among the ornamental cabbages we mention: Brassica oleracea ‘Pigeon white’ with central leaves great and white and external ones green; B. oleracea ‘Songbird white’ with rounded leaves, erect, very elegant, of white colour; B. oleracea ‘Pigeon victoria’ with bicolour leaves lightly wavy; B. oleracea ‘Glamour red’ with elegant intense red leaves.
B. oleracea ‘Crane White’ is suitable to be utilized in the settings and in the seasonal bouquets. B. oleracea var. acephala ‘Osaka’, has curly leaves forming rosettes with the centre of pink, violet, red or white colour that grants a delicious touch of colour to the autumn garden and in early winter.

Lobularia maritima, native to western and central Maditerranean area, is a perennial species, frequent in gardens, that has originated various incredible cultivars, like this, gaudy, with white and violet flowers © Sandy Steinman
Generally, these plants require sunny locations and almost daily waterings, but if cultivated in half shade, every two or three days.
The cultivations begins from the seed, on seedbed, in late spring, the subsequent transplant is to be done with the use of small pots filled with moist loam, that are to be put in luminous places but not in direct sun. It is even possible to buy the seedlings, ready for the use, from the month of August.
The ornamental cabbage is fit to decorate the flower beds of the gardens, balconies and terraces and also for realizing floral compositions.
It loves the locations luminous and sunny even for several hours per day.
It adapts to various types of substrata such as humid soil, well drained to avoid water stagnations. In spring it produces inflorescences with yellow flowers.
Finally, the family includes about one hundred of species of infesting grasses of south-western Asia and of Europe, of which 11 species of Lepidium have become invasive and noxious in North-West America.
The Brassicaceae, particularly the cultivated ones, are subject to adversities caused by various parasites.
Among the cryptogams we mention the alternariosis [Alternaria brassicae (Berk.) Sacc.] that causes dark spots all over the plant.
The clubroot (Plasmodiophora brassicae Woronin) is a pathogen that infects the roots of several species of brassicas thanks to its spores that may resist even many years in the ground even if the host is absent.
The cabbage black spots [Mycosphaerella brassicicola (Duby) Lindau (1897)] can cause the premature defoliation of the plants attacked.
The white rust [Albugo candida (Pers.) Kuntze] is an oomycete attacking the stems, the leaves and the inflorescences and appears as a mass of whitish pustules it uses for the reproduction.
Among the Peronosporaceae we mention the Hyaloperonospora brassicae (Gäum.) Göker, Voglmayr, Riethm., Weiss & Oberw., pathogen attacking all main horticultural species of Brassica. The disease initially manifests with yellow spots, that after the collection, turn black on the upper surface of the leaves and like a white mildew on the lower surface. It severely affects the cauliflower and broccoli causing a premature senescence and favouring the bacterial deterioration in the cold rooms.

Other Lobularia maritima variety with white flowers. The Brassicaceae, apart some weed, are useful plants, alimentary, medicinal and beautiful © Sandy Steinman
The affected Brussels sprouts display black spots that reduce their quality.
The bacterium Xanthomonas campestris pv. campestris (Dowson) Dye, et al. (1980) causes the black rot. Pectobacterium carotovorum (Jones, 1901) Waldee, 1945, is another bacterium that causes the bacterial soft rot to the napa cabbage (Brassica rapa subsp. pekinensis).
Also many insects cause rottenness to the roots and, sometimes, to the petioles of the leaves such as the aphid Myzus persicae (Sulzer, 1776), the larvae of the cabbage moth [i.e. Mamestra brassicae (Linnaeus, 1758)] and the larvae of the cabbage fly [Delia radicum (Linnaeus, 1758)].
The grubs of Pieris rapae (Linnaeus, 1758) and of other butterflies of the family Pieridae are noxious because they nourish of many species of the family, cultivated as well as spontaneous.
Finally, nematodes, rodents and snails alos may cause damage to the cultivated Brassicaceae.

Curious questioning look of a Clypeola jonthlaspi with flowers and fruits. Brassicaceae taxonomy is constantly evolving. Hereunder the 2021 one © Sarah Georg
Genera divided in tribes
Aethionemeae: Aethionema W.T. Aiton Cardamineae: Aplanodes Marais, Armoracia P. Gaertn., B. Mey. & Scherb., Barbarea W.T. Aiton, Bengt-jonsellia Al-Shehbaz, Cardamine L., Iodanthus (Torr. & A. Gray) Steud., Leavenworthia Torr., Nasturtium W.T. Aiton, Ornithocarpa Rose, Planodes Greene, Pteroneurum DC., Rorippa Scop., Selenia Nutt., Sisymbrella Spach
Lepidieae: Cyphocardamum Hedge, Lepidium L., Lithodraba Boelcke
Smelowskieae: Smelowskia C.A. Mey. ex Ledeb.
Alyssopsideae : Alyssopsis Boiss., Calymmatium O.E. Schulz, Dielsiocharis O.E. Schulz, Olimarabidopsis Al-Shehbaz, O’Kane & R.A. Price.
Boechereae: Anelsonia J.F. Macbr. & Payson, Boechera Á. Löve & D. Löve, Borodinia N. Busch, Cusickiella Rollins, Nevada N.H. Holmgren, Phoenicaulis Nutt., Polyctenium Greene, Sandbergia Greene, Yosemitea P.J. Alexander & Windham
Camelineae: Arabidopsis (DC.) Heynh., Camelina Crantz, Capsella Medik., Catolobus (C.A. Mey.) Al-Shehbaz, Chrysochamela (Fenzl) Boiss., Neslia Desv., Noccidium F.K. Mey., Pseudoarabidopsis Al-Shehbaz, O’Kane & R.A. Price, Bursa Boehmer
Crucihimalayeae: Crucihimalaya Al-Shehbaz, O’Kane & R.A. Price, Ladakiella D. German & Al-Shehbaz
Descurainieae: Descurainia Webb & Berthel., Hornungia Rchb., Ianhedgea Al-Shehbaz & O’Kane, Robeschia Hochst. ex O.E. Schulz,
Trichotolinum O.E. Schulz, Tropidocarpum Hook.
Erysimeae: Erysimum L.
Halimolobeae: Exhalimolobos Al-Shehbaz & C.D. Bailey, Halimolobos Tausch, Mancoa Wedd., Pennellia Nieuwl., Sphaerocardamum S. Schauer
Malcolmieae: Malcolmia W.T. Aiton
Microlepidieae: Arabidella O.E. Schulz, Ballantinia Hook. f. ex E.A. Shaw, Blennodia R. Br., Carinavalva Ising, Cuphonotus O.E. Schulz, Drabastrum (F. Muell.) O.E. Schulz, Geococcus J. Drumm. ex Harv., Harmsiodoxa O.E. Schulz, Irenepharsus Hewson, Menkea Lehm.,
Microlepidium F. Muell., Pachycladon Hook. f., Pachymitus O.E. Schulz, Phlegmatospermum O.E. Schulz, Scambopus O.E. Schulz, Stenopetalum Br. ex DC.
Oreophytoneae: Murbeckiella Rothm., Oreophyton O.E. Schulz
Physarieae: Dimorphocarpa Rollins, Dithyrea Harv., Lyrocarpa Hook. & Harv., Nerisyrenia Greene, Paysonia O’Kane & Al-Shehbaz, Physaria
(Nutt. ex Torr. & A. Gray) A. Gray, Synthlipsis A. Gray,
Turritideae: Turritis L.
Yinshanieae: Yinshania Y.C. Ma & Y.Z. Zhao
Alysseae: Acuston Raf., Alyssoides Mill., Alyssum L., Aurinia DC. Berteroa L., Bornmuellera Hausskn., Brachypus Ledeb., Clastopus Bunge ex Boiss.,
Clypeola L., Cuprella Salmerón-Sánchez, Mota & Fuertes, Degenia Hayek, Fibigia Medik., Galitzkya V.V.Botschantz., Hormathophylla Cullen & T.R.Dudley, Irania Hadac & Chrtek, Lepidotrichum Velen. & Bornm., Lutzia Gand., Meniocus Desv., Odontarrhena C.A.Mey., Phyllolepidum Trinajsti,
Physoptychis Boiss, Pterygostemon V.V.Botschantz., Resetnikia Španiel, Al-Shehbaz, D.A.German & Marhold, Takhtajaniella V.E.Avet.
Anastaticeae : Anastatica L., Cithareloma Bunge, Diceratella Boiss., Diceratium Eigia Soják, Eremobium Boiss., Farsetia Turra, Lachnocapsa Balf. f.,
Lobularia Desv., Marcus-kochia Al-Shehbaz, Maresia Pomel, Morettia DC., Notoceras W.T. Aiton, Parolinia Webb
Aphragmeae: Aphragmus Andrz. ex DC.
Arabideae : Abdra Greene, Acirostrum Y.Z. Zhao, Arabis L., Arcyosperma O.E. Schulz, Athysanus Greene, Aubrieta Adans., Baimashania Al-Shehbaz,
Botschantzevia Nabiev, Dendroarabis (C.A. Mey.) D. German & Al-Shehbaz, Draba L., Drabella (DC.) Fourr., Pachyneurum Bunge, Parryodes Jafri,
Pseudodraba Al-Shehbaz, D. German & M.A. Koch, Pseudoturritis Al-Shehbaz, Scapiarabis M.A. Koch, R. Karl, D. German & Al-Shehbaz,
Sinoarabis R. Karl, D. German, M.A. Koch & Al-Shehbaz, Tomostima Raf.
Asteae: Asta Klotzch ex O.E. Schulz
Biscutelleae: Biscutella L., Megadenia Maxim.
Calepineae Calepina Adans., Goldbachia DC., Leiocarpaea (C.A. Mey.) D. German & Al-Shehbaz
Cochlearieae: Cochlearia L., Ionopsidium Rchb.
Coluteocarpeae: Atropatenia F.K. Mey., Callothlaspi F.K. Mey., Eunomia DC., Friedrichkarlmeyeria Tahir Ali & Thines, Ihsanalshehbazia Tahir Ali & Thines, Kotschyella F.K. Mey., Masmenia F.K. Mey., Microthlaspi F.K. Mey., Neurotropis (DC.) F.K. Mey., Noccaea Moench, Pseudosempervivum (Boiss.) Grossh., Thlaspiceras F.K. Mey., Vania F.K. Mey.
Conringieae: Conringia Heist. ex Fabr., Zuvanda (F. Dvořák) Askerova
Cremolobeae: Aimara Salariato & Al-Shehbaz, Cremolobus DC., Menonvillea DC.
Eudemeae: Alshehbazia Salariato & Zuolaga, Aschersoniodoxa Gilg & Muschl., Brayopsis Gilg & Muschl., Dactylocardamum Al-Shehbaz,
Delpinophyton Speg., Delpinophytum Speg., Eudema Humb. & Bonpl., Onuris Phil., Xerodraba Skottsb.
Heliophileae: Cleome, only synonyms, Heliophila L.
Hillielleae: Hilliella (O.E. Schulz) Y.H. Zhang & H.W. Li
Iberideae: Iberis L., Teesdalia W.T. Aiton
Kernereae: Kernera Medik., Rhizobotrya Tausch
Megacarpaeeae: Megacarpaea DC., Pugionium Gaertn.
Notothlaspideae: Notothlaspi Hook. f.
Schizopetaleae: Mathewsia Hook. & Arn., Schizopetalon Sims
Scoliaxoneae: Scoliaxon Payson
Stevenieae: Macropodium W.T. Aiton, Stevenia Fisch
Brassiceae : Ammosperma Hook. f., Bivonaea DC., Brassica L., Cakile Mill., Carrichtera DC., Ceratocnemum Coss. & Balansa, Coincya Porta & Rigo ex Rouy, Cordylocarpus Desf., Crambe L., Crambella Maire, Didesmus Desv., Diplotaxis DC., Douepea Cambess., Enarthrocarpus Labill., Eremophyton Bég., Eruca Mill., Erucaria Gaertn., Erucastrum C. Presl, Fezia Pitt. ex Batt., Foleyola Maire, Fortuynia Shuttelw. ex Boiss., Guiraoa Coss., Hemicrambe Webb, Henophyton Coss. & Durieu, Hirschfeldia Moench, Horwoodia Turrill, Kremeriella Maire, Moricandia DC., Morisia J. Gay, Muricaria Desv., Nasturtiopsis Boiss., Orychophragmus Bunge, Otocarpus Durieu, Physorhynchus Hook., Pseuderucaria (Boiss.) O.E. Schulz, Psychine Desf., Quezeliantha H. Scholz, Quidproquo Greuter & Burdet, Raffenaldia Godr., Raphanus L., Rapistrum Crantz, Rytidocarpus Coss., Savignya DC., Schouwia DC., Sinalliaria X.F. Jin, Y.Y. Zhou & H.W. Zhang, Sinapidendron Lowe, Sinapis L., Succowia Medik., Trachystoma O.E. Schulz, Vella L., Zahora Lemmel & M.Koch, Zilla Forssk.
Isatideae: Chartoloma Bunge, Glastaria Boiss., Isatis L., Myagrum L., Schimpera Hochst. & Steud. ex Schimper, Texiera Jaub. & Spach
Sisymbrieae: Ochthodium DC., Pseudofortuynia Hedge, Sisymbrium L.
Thelypodieae Chaunanthus O.E. Schulz, Chilocardamum O.E. Schulz, Chlorocrambe Rydb., Dictyophragmus O.E. Schulz, Dryopetalon A. Gray, Dryopetalum A. Gray, Englerocharis Muschl., Hesperidanthus (B.L. Rob.) Rydb., Hollermayera O.E. Schulz, Ivania O.E. Schulz, Mostacillastrum O.E. Schulz, Neuontobotrys O.E. Schulz, Petroravenia Al-Shehbaz, Phlebolobium O.E. Schulz, Phravenia Al-Shehbaz & Warwick, Polypsecadium O.E. Schulz, Pringlea T. Anderson ex Hook. f., Romanschulzia O.E. Schulz, Sarcodraba Gilg & Muschl.,Sibara Greene, Stanleya Nutt., Streptanthus Nutt., Terraria T. Hildebrand & Al-Shehbaz, Thelypodiopsis Rydb., Thelypodium Endl., Thysanocarpus Hook., Warea Nutt., Weberbauera Gilg. & Muschl., Zuloagocardamum Salariato & Al-Shehbaz
Eutremeae: Eutrema R. Br.
Thlaspideae: Alliaria Heist. ex Fabr., Didymophysa Boiss., Elburzia Hedge, Graellsia Boiss., Pachyphragma (DC.) Rchb., Parlatoria Boiss., Peltaria Jacq., Peltariopsis (Boiss.) N. Busch, Pseudocamelina (Boiss.) N. Busch, Pseudovesicaria (Boiss.) Rupr., Sobolewskia M. Bieb., Thlaspi L.
Anchonieae: Anchonium DC., Eremoblastus Botsch., Iskandera N. Busch, Matthiola W.T. Aiton, Micrantha F. Dvořák, Microstigma Trautv., Petiniotia Léonard, Sterigmostemum M. Bieb., Synstemon Botsch., Zerdana Boiss.
Buniadeae: Bunias L.
Euclidieae: Anzhenxia Al-Shehbaz & D.A. German, Atelanthera Hook. f. & Thomson, Braya Sternb. & Hoppe, Catenulina Soják, Christolea Cambess., Cryptospora Kar. & Kir., Cymatocarpus O.E. Schulz, Dichasianthus Ovcz. & Yunussov, Dilophia Thomson, Euclidium W.T. Aiton, Lachnoloma Bunge, Leiospora (C.A. Mey.) F. Dvořák, Lepidostemon Hook. f. & Thomson, Leptaleum DC., Metashangrilaia Al-Shehbaz & D.A. German, Neotorularia Hedge & J. Léonard, Octoceras Bunge, Pycnoplinthopsis Jafri, Pycnoplinthus O.E. Schulz, Rhammatophyllum O.E. Schulz, Rudolf–kamelinia Al-Shehbaz & D.A. German, Shangrilaia Al-Shehbaz, J.P. Yue & H. Sun, Sisymbriopsis Botsch. & Tzvelev, Solms-laubachia Muschl., Spryginia Popov, Streptoloma Bunge, Strigosella Boiss., Tetracme Bunge
Hesperideae: Hesperis L., Tchihatchewia Boiss.
Chorisporeae: Chorispora R. Br. ex DC., Diptychocarpus Trautv., Litwinowia Woronow, Parrya R. Br.
Dontostemoneae: Clausia Korn.-Trotsky, Dontostemon Andrz. ex C.A. Mey.
Shehbazieae: Shehbazia D.A. German
Genera not assigned to tribe
Andrzeiowskia Rchb., Asperuginoides Rauschert, Camelinopsis A.G. Mill., Chamira Thunb., Dipoma Franch., Fourraea Greuter & Burdet, Hemilophia Franch., Idahoa A. Nelson & J.F. Macbr., Lunaria L., Parodiodoxa O.E. Schulz, Petrocallis W.T. Aiton, Subularia L.
→ To appreciate the biodiversity within the BRASSICACEAE family please click here.